 Azərbaycanca
Azərbaycanca Беларускі
Беларускі Dansk
Dansk Deutsch
Deutsch Española
Española Français
Français Indonesia
Indonesia Italiana
Italiana 日本語
日本語 Қазақ
Қазақ Lietuvos
Lietuvos Nederlands
Nederlands Português
Português Русский
Русский සිංහල
සිංහල แบบไทย
แบบไทย Türkçe
Türkçe Українська
Українська 中國人
中國人 United State
United State Afrikaans
Afrikaans
Biyosentez, substratların canlı organizmalarda daha karmaşık ürünlere dönüştürüldüğü çok aşamalı, enzim katalizli bir süreçtir. Biyosentezde basit bileşikler modifiye edilir, diğer bileşiklere dönüştürülür veya makromoleküller oluşturmak üzere birleştirilir. Bu süreç genellikle metabolik yollardan oluşur. Bu biyosentetik yollardan bazıları tek bir hücresel organel içinde yer alırken, diğerleri birden fazla hücresel organel içinde yer alan enzimleri içerir. Bu biyosentetik yolların örnekleri arasında çift katlı lipit katmanının bileşenlerinin ve nükleotidlerin üretimi yer alır. Biyosentez genellikle anabolizma ile eş anlamlıdır ve bazı durumlarda birbirinin yerine kullanılır.
Biyosentez olaylarının gerçekleşmesi için öncü bileşikler, kimyasal enerji (örneğin: ATP) ve koenzime sahip katalitik enzimler gereklidir. Bunlar, makromoleküllerin yapı taşları olan monomerleri oluşturur. Bazı önemli makromoleküller, peptit bağları yoluyla birleştirilen ve amino asit monomerlerinden oluşan proteinleri ve fosfodiester bağları yoluyla birleştirilen nükleotitlerden oluşan DNA moleküllerini içermektedir.
Kimyasal reaksiyonların özellikleri
Biyosentez, bir dizi kimyasal reaksiyonlar ile gerçekleşir. Bu reaksiyonların gerçekleşmesi için aşağıdaki özellikler ve unsurlar gereklidir:
- : Bu bileşikler, bir reaksiyondaki başlangıç molekülleri veya substratlarıdır. Bunlar ayrıca belirli bir kimyasal işlemde reaktanlar olarak da görev yapabilirler.
- Kimyasal enerji: Kimyasal enerji, yüksek enerjili moleküller şeklinde bulunabilir. Bu moleküller, enerjik olarak elverişsiz reaksiyonlar için gereklidir. Bu bileşiklerin hidrolizi, bir reaksiyonu ileriye taşır. ATP gibi yüksek enerjili moleküller üç fosfata sahiptir. Çoğu zaman, terminal fosfat hidroliz sırasında ATP'den ayrılır ve başka bir moleküle aktarılır.
- Katalitik enzimler: Bu moleküller, reaksiyonun hızını artırarak ve aktivasyon enerjisini düşürerek bir reaksiyonu katalize eden özel proteinlerdir.
- Kofaktör/Koenzim: Kofaktörler, kimyasal reaksiyonlara yardımcı olan moleküllerdir. Bunlar metal iyonları, NADH ve asetil CoA gibi vitamin türevleri veya ATP gibi vitamin olmayan türevler olabilir.
En basit anlamıyla biyosentezde meydana gelen reaksiyonlar şu formattadır:
Bu temel denklemin ayrıntılı olarak tartışılabilir bazı varyasyonları şunlardır:
- Genellikle çok aşamalı bir reaksiyon yolunun parçası olarak diğer bileşiklere dönüştürülen basit bileşikler. Bu tip reaksiyonun iki örneği, nükleik asitlerin oluşumunda ve translasyondan önce tRNA'nın yüklenmesi sırasında meydana gelir. Bu adımlardan bazıları için kimyasal enerji gereklidir:
- Kofaktörlerin yardımıyla başka bileşiklere dönüştürülen basit bileşikler. Örneğin, fosfolipitlerin sentezi asetil CoA'yı gerektirirken, başka bir zar bileşeni olan sfingolipidlerin sentezi, sfingozin omurgasının oluşumu için NADH ve FADH'yi gerektirir. Bu örnekler için genel denklem şu şekildedir:
- Bir makromolekül oluşturmak için birleşen basit bileşikler. Örneğin, yağ asitleri birleşerek fosfolipitleri oluşturur. Buna karşılık, fosfolipidler ve kolesterol, lipid çift tabakasını oluşturmak için kovalent olmayan bağlarla etkileşime girer. Bu reaksiyon aşağıdaki gibi gösterilebilir:
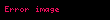
![{\displaystyle {\ce {{{\ddot {O}}nc{\ddot {u}}\ molek{\ddot {u}}l}+Kofakt{\ddot {o}}r->[][enzim]makromolek{\ddot {u}}l}}}](https://www.wikipedia.tr-tr.nina.az/image/aHR0cHM6Ly93d3cud2lraXBlZGlhLnRyLXRyLm5pbmEuYXovaW1hZ2UvYUhSMGNITTZMeTkzYVd0cGJXVmthV0V1YjNKbkwyRndhUzl5WlhOMFgzWXhMMjFsWkdsaEwyMWhkR2d2Y21WdVpHVnlMM04yWnk4ME56WTBNamMzTVRJNU1qRmtaREEzWWpoalpEVTRZV0ptWmpJelpXSXdORFkwT0RCak4yRTEuc3Zn.svg)
Lipit

Birçok karmaşık makromolekül, basit, tekrarlanan yapıların bir modelinde sentezlenir. Örneğin, lipitlerin en basit yapıları yağ asitleridir. Yağ asitleri hidrokarbon türevleridir; bir karboksil grubu "baş" ve bir hidrokarbon zinciri "kuyruk" içerir. Bu yağ asitleri daha büyük bileşenler oluşturur ve bunlar da lipit çift tabakasını oluşturmak için kovalent olmayan etkileşimleri içerir. Yağ asidi zincirleri, membran lipidlerinin iki ana bileşeni olan fosfolipitler ve sfingolipidlerde bulunur. Üçüncü bir ana zar bileşeni olan kolesterol, bu yağ asidi birimlerini içermez.
Fosfolipitler
Tüm biyomembranların temeli, iki katmanlı bir fosfolipit yapısından oluşur. Fosfolipid molekülü amfifildir; hidrofilik bir kutup başı ve hidrofobik kutupsuz bir kuyruk içerir. Fosfolipid kafaları birbirleriyle ve sulu ortamla etkileşime girerken, hidrokarbon kuyrukları kendilerini merkezde sudan uzağa yönlendirir. Bu son etkileşimler, iyonlar ve moleküller için bir bariyer görevi gören çift katmanlı yapıyı yönlendirir.
Çeşitli fosfolipid türleri vardır ancak sentez yolları farklıdır. Fosfolipid sentezinde ilk adım, endoplazmik retikulumda ve mitokondride gerçekleşir. Sentez sırasında veya da oluşur. Sentez şu şekilde gerçekleşmektedir:

Sentez, tarafından sağlanan bir yağ asidi zincirinin eklenmesi yoluyla lizofosfatidata dönüştürülen gliserol 3-fosfat ile başlar. Daha sonra, lizofosfatidat, ikinci bir açil CoA'nın katkıda bulunduğu başka bir yağ asidi zincirinin eklenmesi yoluyla fosfatidata dönüştürülür; bu adımların tümü, gliserol fosfat enzimi tarafından katalize edilir. Fosfolipid sentezi endoplazmik retikulumda devam eder ve biyosentez, belirli fosfolipidin bileşenlerine bağlı olarak farklılık gösterir.
Sfingolipitler
Fosfolipidler gibi, bu yağ asidi türevlerinin de bir kutup başı ve kutupsal olmayan kuyrukları vardır. Fosfolipidlerin aksine, sfingolipidlerin bir sfingozin omurgası vardır. Sfingolipidler ökaryotik hücrelerde, özellikle merkezi sinir sisteminde bol miktarda bulunur. Örneğin, sfingomyelin sinir lifleri, miyelin kılıfının bir parçasıdır.
Sfingolipidler, bir sfingozin omurgasının amino grubuna bağlı bir yağ asidi zincirinden oluşan seramidlerden oluşur. Bu seramidler, sfingosinin asilasyonundan sentezlenir. Sfingosinin biyosentezi şu şekildedir:

Sfingozin sentezi sırasında ve serin, dehidrosfingosin oluşumuyla sonuçlanan bir kondenzasyon reaksiyonuna girer. Bu ürün daha sonra FAD tarafından oksidasyon reaksiyonu yoluyla sfingosine dönüştürülen dihidrospingozini oluşturmak üzere indirgenir.
Kolesterol
Kolesterol, sterol adı verilen bir molekül sınıfına aittir. Sterollerin dört kaynaşmış halkası ve bir hidroksil grubu vardır. Kolesterol özellikle önemli bir moleküldür. Sadece lipid zarlarının bir bileşeni olarak görev yapmakla kalmaz, aynı zamanda kortizol, testosteron ve östrojen de dahil olmak üzere çeşitli steroid hormonlarının yapısına katılır.
Kolesterol, asetil-CoA'dan sentezlenir. Sentezi şu şekildedir:

Daha genel olarak, bu sentez üç aşamada gerçekleşir. Birinci aşama sitoplazmada gerçekleşir ve ikinci ve üçüncü aşama endoplazmik retikulumda meydana gelir. Aşamaları ise şu şekildedir:
- Kolesterolün yapı taşı olan sentezi
- Altı molekül izopentenil fosfatın yoğunlaşmasıyla skualen oluşumu
- Birkaç enzimatik reaksiyon yoluyla skualenin kolesterole dönüşümü
Nükleotit
Nükleotitlerin biyosentezi, substratları daha karmaşık ürünlere dönüştüren enzim katalizli reaksiyonlardan oluşmaktadır. Nükleotitler, DNA ve RNA'nın yapı taşlarıdır. Nükleotitler, RNA'daki riboz şekerinden ve DNA'daki deoksiriboz şekerden oluşan beş üyeli bir halkadan oluşur; bu şekerler bir glikosidik bağ ve bir fosfat grubu ile bir pürin veya pirimidin bazına bağlanır.
Pürinler

DNA nükleotidlerinden adenozin ve , bir glikosidik bağ ile bir riboz şekerine bağlı bir pürin bazından oluşur. RNA nükleotidleri deoksiadenozin ve deoksiguanozin durumunda, pürin bazları bir glikozidik bağ ile bir deoksiriboz şekere bağlanır. DNA ve RNA nükleotidlerindeki pürin bazları, çoğu tek hücreli organizmada bulunan on iki aşamalı bir reaksiyon mekanizmasında sentezlenir. Daha gelişmiş ökaryotlar, on reaksiyon adımında benzer bir reaksiyon mekanizması kullanır. Pürin bazları, fosforibosil pirofosfatın (PRPP), pürin baz biyosentezindeki ilk anahtar ara madde olan inozin monofosfata (IMP) dönüştürülmesiyle sentezlenir. IMP'nin daha fazla enzimatik modifikasyonu, nükleotitlerin adenozin ve guanozin bazlarını üretir. Pürinlerin biyosentezi şu şekilde gerçekleşir:
- Pürin biyosentezindeki ilk adım, tarafından gerçekleştirilen bir yoğunlaşma reaksiyonudur. Bu enzim, amino grubunu glutaminden PRPP'ye aktararak oluşturur. Aşağıdaki adım, ATP'den bir fosfat grubunun eklenmesiyle glisinin aktivasyonunu gerektirir.
- GAR sentetaz, aktifleştirilmiş glisinin PRPP üzerinde yoğunlaştırılmasını gerçekleştirerek glisinamid ribonükleotid (GAR) oluşturur.
- GAR transformilaz, GAR'ın amino grubuna bir formil grubu ekleyerek formilglisinamid ribonükleotid (FGAR) oluşturur.
- FGAR amidotransferaz, bir nitrojen grubunun FGAR'a eklenmesini katalize ederek formilglisinamidin ribonükleotid (FGAM) oluşturur.
- FGAM siklaz, 5'li halkasını 5-aminoimidazol ribonükleotidi (AIR) oluşturan bir su molekülünün çıkarılmasını içeren halka kapanmasını katalize eder.
- N5-CAIR sentetaz, bir karboksil grubunu transfer ederek ara ürün olan N5-karboksiaminoimidazol ribonükleotidi (N5-CAIR) oluşturur.
- N5-CAIR mutazı, karboksil fonksiyonel grubunu yeniden düzenler ve onu imidazol halkasına aktararak karboksiamino-imidazol ribonükleotidi (CAIR) oluşturur. AIR'den CAIR oluşumunun iki aşamalı mekanizması çoğunlukla tek hücreli organizmalarda bulunur. Daha yüksek ökaryotlar, bir karboksil grubunu doğrudan AIR imidazol halkasına aktaran ve CAIR oluşturan AIR karboksilaz enzimini içerir.
- SAICAR sentetaz, aspartat ile imidazol halkasının eklenen karboksil grubu arasında, (SAICAR) oluşturan bir peptit bağı oluşturur.
- SAICAR liyaz, eklenen aspartatın karbon iskeletini çıkararak amino grubunu terk eder ve (AICAR) oluşturur.
- AICAR transformilaz, bir karbonil grubunu AICAR'a aktararak (FAICAR) oluşturur.
- Son adım, pürin halkasının kapanmasını gerçekleştiren ve inozin monofosfat ara ürününü oluşturan IMP sentaz enzimi ile gerçekleşir.
Pirimidinler

Bir glikozidik bağ yoluyla riboz şekerine bağlanan diğer DNA ve RNA nükleotid bazları timin, sitozin ve urasildir (Urasil RNA'ya özgüdür). (UMP) biyosentezinde mitokondriyal iç zarda bulunan bir enzim ve sitozolde bulunan çok işlevli enzimler bulunmaktadır. Pirimdinlerin biyosentezi şu şekilde gerçekleşir:
- İlk adımda, karbamoil fosfat oluşturmak için ATP'ye bağlı bir reaksiyonda glutamin ile CO2'yi birleştiren enzim bulunmaktadır.
- , üridosüksinat oluşturmak için karbamoil fosfatı aspartat ile yoğunlaştırır .
- , oluşturmak için su kaybeden bir reaksiyon olan halka kapanmasını gerçekleştirir.
- Mitokondriyal iç zar içinde yer alan dihidroorotatı oksitler.
- Orotat, fosforibosil hidrolaz (OMP pirofosforilaz), orotatı PRPP ile yoğunlaştırarak orotidin-5'-fosfat oluşturur.
- , orotidin-5'fosfatın UMP'ye dönüşümünü katalize eder.
Üridin nükleotid bazı sentezlendikten sonra diğer bazlar olan sitozin ve timin sentezlenir. Sitozin biyosentezi, UMP'nin 'ye dönüşümünü içeren iki aşamalı bir reaksiyondur. UMP'ye fosfat ilavesi, bir kinaz enzimi tarafından katalize edilir. enzimi bir sonraki reaksiyon adımını katalize eder. bir amino grubunu glutaminden üridine transfer ederek UTP'nin 'ye dönüştürülmesi, CTP'nin sitozin bazını oluşturur.
UTP + ATP + glutamin ⇔ CTP + ADP + glutamat reaksiyonu şu şekildedir:

Sitozin, hem DNA hem de RNA'da bulunan bir nükleotittir. Ancak urasil sadece RNA'da bulunur. Bu nedenle UTP sentezlendikten sonra DNA'ya dahil edilmesi için formuna dönüştürülmesi gerekir. Bu dönüşüm, enzimi ile gerçekleşir. Deoksiriboz oluşturmak için riboz şekerinin 2 -OH'sini uzaklaştıran bu reaksiyon, şekere bağlı bazlardan etkilenmez. Bu özgül olmama, ribonükleozit trifosfat redüktazın benzer bir mekanizma ile tüm nükleotid trifosfatları deoksiribonükleotite dönüştürmesine izin verir.
Urasilin aksine, timin bazları çoğunlukla RNA'da değil DNA'da bulunur. Hücreler normalde RNA'daki riboz şekerlerine bağlı timin bazları içermez, bu da hücrelerin yalnızca deoksiriboza bağlı timini sentezlediğini gösterir. Timidilat sentetaz enzimi, timin kalıntılarının dUMP'den dTMP'ye sentezlenmesinden sorumludur. Bu reaksiyon, dTMP oluşturmak için bir metil grubunu dUMP'nin urasil bazına aktarır.
Timidilat sentaz reaksiyonu şu şekildedir: dUMP + 5,10-metilentetrahidrofolat ⇔ dTMP + dihidrofolat
DNA

Ökaryotik ve prokaryotik canlılarda DNA sentezi arasında farklılıklar olmasına rağmen aşağıdaki bölümler her iki organizma tarafından paylaşılan DNA replikasyonunun temel özelliklerini anlatmaktadır:
DNA, fosfodiester bağları ile birleştirilen nükleotitlerden oluşur. Çekirdekte gerçekleşen DNA sentezi, yarı koruyucu bir süreçtir. Bu, elde edilen DNA molekülünün ana yapıdan orijinal bir iplik ve yeni bir iplik içerdiği anlamına gelir. DNA sentezi, dört deoksinükleosit trifosfat, bir şablon iplik ve nükleotitlerin dahil edileceği serbest 3'OH'li bir primer gerektiren bir DNA polimeraz ailesi tarafından katalize edilir.
DNA replikasyonunun gerçekleşmesi için, DNA sarmalını çözen helikaz adı verilen enzimler tarafından bir replikasyon çatalı oluşturulur. Replikasyon çatalındaki topoizomerazlar, DNA'nın çözülmesinin neden olduğu süper bobinleri kaldırır ve tek iplikli DNA bağlayıcı proteinler, replikasyondan önce stabilize edilmiş iki tek iplikli DNA şablonunu korur.
DNA sentezi, serbest bir 3'OH ile bir RNA primeri yapan RNA polimeraz primeri tarafından başlatılır. Bu primer, tek iplikli DNA şablonuna bağlıdır ve DNA polimeraz, nükleotidleri dahil ederek zinciri uzatır; DNA polimeraz ayrıca yeni sentezlenen DNA zincirini de düzeltir.
DNA polimeraz tarafından katalize edilen polimerizasyon reaksiyonu sırasında, büyüyen zincirin 3'OH'si tarafından bir deoksinükleosit trifosfatın en içteki fosfor atomu üzerinde bir nükleofilik saldırı meydana gelir; bu, yeni bir nükleotidi bağlayan ve pirofosfat salan bir fosfodiester bağının oluşumunu sağlar.
Replikasyon sırasında aynı anda, sürekli sentezlenen ve replikasyon çatalına doğru büyüyen öncü iplik ve Okazaki parçalarında süreksiz olarak yapılan ve replikasyon çatalından uzaklaşan gecikmeli iplik olmak üzere iki tip iplik oluşur. Okazaki parçaları, sürekli bir iplik oluşturmak için DNA ligaz ile ve kovalent bağ ile birleştirilir. Daha sonra DNA replikasyonunu tamamlamak için RNA primerleri çıkarılır ve oluşan boşluklar DNA ile değiştirilir ve DNA ligaz yoluyla birleştirilir.
Amino asit
Protein, peptid bağlarıyla birbirine bağlanan amino asitlerden oluşan bir polimerdir. Doğada bulunan 300'den fazla amino asit vardır. Bunlardan sadece yirmi tanesi standart/temel amino asitler olarak bilinir ve proteinin yapı taşlarıdır. Sadece yeşil bitkiler ve çoğu mikrop, tüm canlı türlerinin ihtiyaç duyduğu 20 standart amino asidin tamamını sentezleyebilir. Memeliler, yirmi standart amino asitten yalnızca onunu sentezleyebilir. Diğer amino asitler olan valin, metionin, lösin, izolösin, fenilalanin, lizin, treonin ve triptofan, histidin ve arginin beslenme yoluyla elde edilir.
Amino asidin temel yapısı

Standart amino asitlerin genel yapısı, bir birincil amino grubu, bir karboksil grubu ve bağlı fonksiyonel grubu içerir. Amino asitler, fonksiyonel gruplar nedeniyle birbirinden ayrılır. α-karbona bağlı üç farklı grubun bir sonucu olarak, amino asitler asimetrik moleküllerdir. Glisin hariç tüm standart amino asitler için α-karbon bir kiral merkezdir. Glisin durumunda, a-karbon iki hidrojen atomuna sahiptir, bu durum moleküle simetri ekler. Prolin hariç, yaşamda bulunan tüm amino asitler L-izoform konformasyonuna sahiptir. Prolin, amino grubu ile bir halka oluşturan α-karbon üzerinde fonksiyonel bir gruba sahiptir.

Amino asitlerin azot kaynağı
Amino asit biyosentezindeki önemli bir adım, a-karbon üzerine bir azot grubunun dahil edilmesidir. Hücrelerde, azot gruplarını dahil etmenin iki ana yolu vardır. Birinci yol, glutaminin amit amino grubunu uzaklaştıran ve onu üzerine transfer eden ve iki glutamat molekülü üreten (GOGAT) enzimi ile dahil etmektir. Bu kataliz reaksiyonunda azot kaynağı olarak glutamin görev yapar. Bu reaksiyon yukarıdaki resimde gösterilmiştir.
Amino asitlerin a-karbonuna azot dahil etmenin diğer yolu, (GDH) enzimi ile dahil etmektir. GDH, amonyağı 2-oksoglutarat üzerine aktarabilir ve glutamat oluşturabilir. Ayrıca, (GS) enzimi, amonyağı glutamata aktarabilir ve glutamini yenileyerek glutamin sentezleyebilir.
Amino asitlerin glutamat ailesi
Amino asitlerin ailesi, amino asit glutamattan türetilen amino asitleri içerir. Bu aile glutamat, glutamin, prolin ve argininden oluşmaktadır. Bu aile ayrıca türetilen amino asit lizini de içerir.
Glutamat ve glutamin biyosentezi, azot asimilasyonunda önemli bir adımdır. GOGAT ve GDH enzimleri, azot asimilasyon reaksiyonlarını katalize eder.
Glutamat ve glutamin biyosentezi, azot asimilasyonunda önemli bir adımdır. GOGAT ve GDH enzimleri, azot asimilasyon reaksiyonlarını katalize eder.
Bakterilerde enzimi, bir fosfat grubunu ATP'den glutamata aktararak prolin biyosentezini başlatır. Sonraki reaksiyon, L-glutamat 5-fosfatın grubunun indirgenmesini katalize eden enzim (P5CS) tarafından katalize edilir. Bu, kendiliğinden pirolin-5-karboksilata siklize olan glutamat semialdehit oluşumu ile sonuçlanır. Pirolin-5-karboksilat, bir prolin amino asidi verecek şekilde pirolin-5-karboksilat redüktaz (P5CR) enzimi tarafından daha da indirgenir.
Bakterilerde arginin biyosentezinin ilk adımında, asetil grubunun N-α pozisyonunda asetil-CoA'dan aktarılmasıyla glutamat asetillenir; bu kendiliğinden siklizasyonu önler. N-asetilglutamat sentaz enzimi (glutamat N-asetiltransferaz), asetilasyon adımını katalize etmekten sorumludur. Sonraki adımlar, ve asetilornitin/süksinildiamino pimelat aminotransferaz enzimleri tarafından katalize edilir ve N-asetil-L-ornitin verir. Asetillornitin asetil grubu, (AO) veya (OAT) enzimi tarafından çıkarılır ve bu, verir. Daha sonra ve enzimleri ornitini arginine dönüştürür.

İki farklı lizin biyosentetik yolu vardır. Diaminopimelik asit yolu ve a-aminoadipat yolu olmak üzere ikiye ayrılır. İki sentetik yoldan en yaygın olanı ; lizin elde etmek için aspartata karbon grupları ekleyen birkaç enzimatik reaksiyondan oluşur:
- Aspartat kinaz, aspartatı fosforile ederek ve aspartil fosfat üreterek diaminopimelik asit yolunu başlatır.
- Aspartat semialdehit dehidrojenaz, aspartil fosfatın NADPH'ye bağlı indirgenmesini katalize ederek aspartat semialdehit verir.
- 4-hidroksi-tetrahidrodipikolinat sentaz, β-aspartil-4-semialdehite bir piruvat grubu ekler ve bir su molekülü çıkarılır. Bu neden olur ve (2S,4S)-4-hidroksi-2,3,4,5-tetrahidrodipikolinata yol açar.
- 4-hidroksi-tetrahidrodipikolinat redüktaz, (2S,4S)-4-hidroksi-2,3,4,5-tetrahidrodipikolinatın NADPH tarafından indirgenmesini katalize ederek Δ'-piperidein-2,6-dikarboksilat (2,3,4, 5 -tetrahidrodipikolinat) ve H20 ortaya çıkarır.
- Tetrahidrodipikolinat asiltransferaz, halka açılmasıyla sonuçlanan asetilasyon reaksiyonunu katalize eder ve N-asetil a-amino-ε-ketopimelat verir.
- , N-asetil a-amino-ε-ketopimelat'ın keto grubunu uzaklaştıran ve N-süksinil-L-diaminopimelat verecek şekilde bir amino grubuyla değiştiren transaminasyon reaksiyonunu katalize eder.
- N-asildiaminopimelat deasilaz, L,L-diaminopimelat verecek şekilde N-süksinil-L-diaminopimelatın deasilasyonunu katalize eder.
- , L,L-diaminopimelatın L,L-diaminopimelatın mezo formuna dönüşümünü katalize eder.
- , karboksil grubunun çıkarılmasını katalize ederek L-lisin verir.
Amino asitlerin serin ailesi
Amino asidin serin ailesi; serin, sistein ve glisinden oluşmaktadır. Çoğu mikroorganizma ve bitki, amino asit sisteininden metiyonini sentezlemek için kükürt elde eder. Ayrıca serinin glisine dönüşümü, metiyonin ve histidinin biyosentezi için gerekli karbonları sağlar.
Serin biyosentezi sırasında, fosfogliserat dehidrojenaz enzimi, 3-fosfo-D-gliseratı oksitleyerek veren ilk reaksiyonu katalize eder. Reaksiyonda, bir amino grubunu glutamattan 3-fosfonooksipiruvat üzerine transfer etirilerek L-fosfoserin veren enzim tarafından katalize edilir. Son adımda, L-serin verecek şekilde L-fosfoserin'i fosforile eden enzim tarafından katalize edilir.
Glisin biyosentezi için bilinen iki yol vardır: Bir yolunda ana karbon kaynağı olarak etanol ve asetat kullanan organizmalar, glisin sentezlemek için yolu kullanır. Glisin biyosentezinin diğer yolu, glikolitik yol olarak bilinir. Bu yol, glikolizin ara ürünlerinden sentezlenen serini glisine dönüştürür. Glikolitik yolda, enzimi, glisin elde etmek için serinin bölünmesini katalize eder ve bölünmüş serin karbon grubunu aktararak oluşturur.
Sistein biyosentezi, inorganik kükürtün dahil olduğu, iki aşamalı bir reaksiyondur. Mikroorganizmalarda ve bitkilerde, enzimi, asetil grubunun asetil-CoA'dan L-serine transferini katalize ederek O-asetil-L-serin verir. enzimi tarafından katalize edilen reaksiyon adımı, sistein verecek şekilde O-asetil-L-serinin asetil grubunu sülfür ile değiştirir.
Aspartat amino asit ailesi
Aspartat amino asit ailesi; treonin, lizin, metionin, izolösin ve aspartattan oluşmaktadır. Lizin ve izolösin, karbon iskeletlerinin bir kısmı piruvattan türetilmiş olsa da, aspartat ailesinin bir parçası olarak kabul edilir. Metiyonin durumunda, metil karbon serin ve kükürt grubundan türetilir, ancak çoğu organizmada sisteinden türetilir.
Aspartat biyosentezi, tek bir enzim tarafından katalize edilen tek adımlı bir reaksiyondur. Aspartat aminotransferaz enzimi, bir amino grubunun aspartattan a-ketoglutarat üzerine transferini katalize ederek glutamat ve oksaloasetat verir. Asparagin, aspartat üzerine bir amino grubunun ATP'ye bağımlı eklenmesiyle sentezlenir; asparagin sentetaz, asparagin verecek şekilde glutamin veya çözünür amonyaktan aspartata azot eklenmesini katalize eder.

Lizinin diaminopimelik asit biyosentetik yolu, aspartat amino asit ailesine aittir. Bu yol, aspartatı lizine dönüştüren dokuz enzim katalizli reaksiyonu içerir.
- Aspartat kinaz, ATP'den aspartil-β-fosfat veren karboksilat grubuna ATP'den bir fosfor aktararak diaminopimelik asit yolundaki ilk adımı katalize eder.
- Aspartat-semialdehit dehidrojenaz, aspartat-β-semialdehit verecek şekilde aspartil-β-fosfatın fosforilasyonu yoluyla indirgeme reaksiyonunu katalize eder.
- Dihidrodipikolinat sentaz, aspartat-β-semialdehitin piruvat ile katalize ederek dihidrodipikolinik asit verir.
- , dihidrodipikolinik asidin indirgenmesini katalize ederek tetrahidrodipikolinik asit verir.
- , bir süksinil grubunun süksinil-CoA'dan tetrahidrodipikolinik asit üzerine transferini katalize ederek N-süksinil-L-2,6-diaminoheptandioat verir.
- N-süksinildiaminopimelat aminotransferaz, bir amino grubunun glutamattan N-süksinil-L-2,6-diaminoheptandioat üzerine transferini katalize ederek N-süksinil-L,L-diaminopimelik asit verir.
- , asil grubunun N-süksinil-L,L-diaminopimelik asitten çıkarılmasını katalize ederek L,L-diaminopimelik asit verir.
- verecek şekilde L,L-diaminopimelik asidin a-karbonunun ters çevrilmesini katalize eder.
- Siaminopimelat dekarboksilaz, karbon dioksit grubunu mezo-diaminopimelik asitten uzaklaştırarak L-lizin veren lizin biyosentezindeki son adımı katalize eder.
Proteinler


Protein sentezi, translasyon adı verilen bir süreçle gerçekleşir. Translasyon sırasında, mRNA adı verilen genetik materyal, bir protein polipeptit zinciri oluşturmak için ribozomlar tarafından okunur. Bu işlem, bir ucunda amino asitleri bağlayarak ve diğer ucunda mRNA ile etkileşime girerek bir adaptör görevi gören transfer RNA'sını (tRNA) gerektirir; tRNA ve mRNA arasındaki ikinci eşleşme, zincire doğru amino asidin eklenmesini sağlar. Protein sentezi; başlama, uzama ve sonlandırma olmak üzere üç aşamada gerçekleşir. Prokaryotik canlılarda translasyon, farklıdır.
Translasyon öncesi
Translasyon başlamadan önce, belirli bir amino asidi karşılık gelen tRNA'ya bağlama işlemi gerçekleşmelidir. tRNA yüklemesi olarak adlandırılan bu reaksiyon, tarafından katalize edilir. Belirli bir amino asidin tanınmasından ve yüklenmesinden belirli bir tRNA sentetaz sorumludur. Ayrıca, bu enzim, tRNA ile aynı kökenli amino asidi arasında doğru bağlanmayı sağlamak için özel ayırıcı bölgelere sahiptir. Bir amino asidi karşılık gelen tRNA'ya bağlamanın ilk adımı, aminoasil-AMP'nin oluşumudur:

Bunu aminoasil grubunun aminoasil-AMP'den bir tRNA molekülüne transferi izler. Ortaya çıkan molekül aminoasil-tRNA'dır:
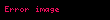
Aminoasil tRNA sentetaz tarafından katalize edilen bu iki adımın kombinasyonu, büyüyen polipeptit zincirine amino asitler eklemeye hazır olan yüklü bir tRNA üretir.
Bir amino asidi bağlamaya ek olarak, tRNA, mRNA'da da bulunan kodon adı verilen spesifik nükleotit üçlüleri ile baz çiftleri oluşturan, antikodon adı verilen üç nükleotit birimine sahiptir; kodonlar belirli bir amino asidi kodlar. Bu etkileşim, protein sentezi için bölge görevi gören ribozom sayesinde mümkündür. Ribozom, aminoasil bölgesi (A bölgesi), peptidil bölgesi (P bölgesi) ve çıkış bölgesi (E bölgesi) olmak üzere üç tRNA bağlanma bölgesine sahiptir.
Bir mRNA transkriptinde çok sayıda kodon vardır. Bu nedenle bir amino asidin birden fazla kodon ile etkileşimi çok yaygındır; buna dejenerasyon denir. Toplamda, 20 amino asitten biri için her kodun 61'i olmak üzere 64 kodon vardır, geri kalan kodonlar ise zincir sonlandırmasını belirtir.
Translasyonun aşamaları
Translasyonun başlatma, uzama ve sonlandırma olmak üzere üç aşaması vardır.

1. Adım: Başlatma
Başlatma aşamasının tamamlanması için bu üç olayın gerçekleşmesi gerekmektedir.
- Ribozomun mRNA'ya alınması
- Yüklü bir başlatıcı tRNA'nın ribozomun P bölgesine bağlanması
- Ribozomun mRNA'nın başlangıç kodonu ile doğru hizalanması
2. Adım: Uzama
Başlatmayı takiben, polipeptit zinciri, antikodon:kodon etkileşimleri yoluyla uzatılır ve ribozom, polipeptit zincirine birer birer amino asitler ekler. Amino asitlerin doğru eklenmesini sağlamak için aşağıdaki adımlar gerçekleşmelidir:
- Doğru tRNA'nın ribozomun A bölgesine bağlanması
- A bölgesindeki tRNA ile P bölgesindeki tRNA'ya bağlı polipeptit zinciri arasında peptit bağının oluşması
- Üç nükleotid tarafından tRNA-mRNA kompleksinin translokasyonu veya ilerlemesi
Translokasyon, E bölgesindeki tRNA'nın hareketini başlatır ve tRNA'yı A bölgesinden P bölgesine kaydırır, A bölgesini gelen bir tRNA'nın başka bir amino asit eklemesi için serbest bırakır.
3. Adım: Sonlandırma
Translasyonun son aşaması, bir durdurma kodonu A bölgesine girdiğinde gerçekleşir. Ardından, aşağıdaki adımlar gerçekleşir:
- P bölgesinde bulunan tRNA'dan polipeptit zincirinin hidrolizine neden olan salım faktörleri tarafından kodonların tanınması
- Polipeptit zincirinin salınımı
- Gelecekte gerçekleşecek translasyon süreçleri için ribozomun ayrılması ve geri dönüşümü
Translasyonda görev alan önemli moleküller:
| Molekül | Bulunduğu adım | Görevi |
|---|---|---|
| tRNA sentetaz | Başlatmadan önce | tRNA şarjından sorumludur. |
| mRNA | Başlatma, uzama, sonlandırma | Protein sentezi için şablon ve amino asitleri kodlayan kodonlar olarak adlandırılan bölgeleri bulundurur. |
| tRNA | A, P, E ribozom bölgelerine bağlanır; doğru amino asidin büyüyen polipeptit zincirine dahil edilmesini sağlamak için mRNA kodonu ile antikodon baz çiftlerini barındırır. | |
| Ribozom | Protein sentezini yönetir ve peptit bağı oluşumunu katalize eder. |
Makromolekül eksikliği ile iişkili hastalıklar

Biyosentetik yollardaki hatalar, makromoleküllerin malformasyonu veya fonksiyonel moleküllerin yetersiz üretimi dahil olmak üzere zararlı sonuçlara sahip olabilir.
- : Bu bozukluk, LDL için fonksiyonel reseptörlerin yokluğu ile karakterizedir. LDL reseptörlerinin oluşumundaki eksiklikler, endositik yolu bozan, LDL'nin karaciğere ve diğer hücrelere girişini engelleyen hatalı reseptörlere neden olabilir. Bu, kan plazmasında LDL birikmesine neden olur, bu da arterleri daraltan ve kalp krizi riskini artıran aterosklerotik plaklara neden olur.
- : Bu genetik hastalık kendini yaralama, zihinsel yetersizlik ve gut ile karakterizedir. Pürin nükleotid oluşumu için gerekli bir enzim olan yokluğundan kaynaklanır. Enzim eksikliği, gerekli nükleotidlerin seviyesini azaltır ve biyosentez ara ürünlerinin birikmesine neden olur, bu da yukarıda belirtilen olağandışı davranışlarla sonuçlanır.
- (SCID): SCID, T hücrelerinin kaybı ile karakterizedir. Bu bağışıklık sistemi bileşenlerinin eksikliği, etkilenen bireyler immünolojik hafıza geliştiremediğinden, bulaşıcı ajanlara duyarlılığı arttırır. Bu immünolojik bozukluk, dATP birikmesine neden olan aktivitesindeki bir eksiklikten kaynaklanır. Bu dATP molekülleri daha sonra DNA sentezini önleyen ribonükleotid redüktazı inhibe eder.
- Huntington hastalığı: Bu nörolojik hastalık, DNA sentezi sırasında meydana gelen hatalardan kaynaklanır. Bu hatalar veya mutasyonlar, gendeki genişleyen CAG trinükleotit tekrarları tarafından kodlanan tekrarlayan glutamin tortuları içeren bir mutant Huntingtin proteininin ekspresyonuna yol açar. Huntington hastalığı, nöronal kayıp ve gliozis ile karakterizedir. Hastalığın belirtileri; hareket bozukluğu, bilişsel gerileme ve davranış bozukluğudur.
Ayrıca bakınız
Kaynakça
- ^ a b c d Alberts, Bruce (2008). Molecular Biology of the Cell (5. bas.). New York: Garland Science. ISBN .
- ^ Zumdahl, Steven S. Zumdahl, Susan A. (2008). Chemistry (8. bas.). CA: Cengage Learning. ISBN .
- ^ Voet, Donald; Voet, Judith G.; Pratt, Charlotte W. (2013). Fundamentals of Biochemistry: Life at the Molecular Level. 4th. Hoboken, NJ: Wiley. ISBN .
- ^ a b c d e Lodish, Harvey (2007). Molecular cell biology. 6. New York: W.H. Freeman. ISBN .
- ^ a b c d e Cox, David L. Nelson, Michael M. (2008). Lehninger principles of biochemistry (5. bas.). New York: W.H. Freeman. ISBN .
- ^ Hanin, Israel (2013). Phospholipids: Biochemical, Pharmaceutical, and Analytical Considerations. Springer. ISBN .
- ^ a b c d Vance, Dennis E.; Vance, Jean E. (2008). Biochemistry of lipids, lipoproteins and membranes (5. bas.). Amsterdam: Elsevier. ISBN .
- ^ Katsaras, J. (2001). Lipid bilayers : structure and interactions ; with 6 tables. Berlinisbn=978-3540675556: Springer.
- ^ a b c d e Stryer, Jeremy M. Berg; John L. Tymoczko; Lubert (2007). Biochemistry (6. bas.). New York: Freeman. ISBN .
- ^ Gault, CR; LM Obeid; YA Hannun (2010). An Overview of sphingolipid metabolism: from synthesis to breakdown. Advances in Experimental Medicine and Biology. 688. s. 1–23. doi:10.1007/978-1-4419-6741-1_1. ISBN . (PMC) 3069696 $2. (PMID) 20919643.
- ^ a b Siegel, George J. (1999). Basic neurochemistry : molecular, cellular and medical aspects (6. bas.). Philadelphia, Pa. [u.a.]: Lippincott Williams & Wilkins. ISBN .
- ^ a b c Harris, J. Robin (2010). Cholesterol binding and cholesterol transport proteins : structure and function in health and disease. Dordrecht: Springer. ISBN .
- ^ a b c d e f Watson, James D. (2007). Molecular biology of the gene (6. bas.). San Francisco, Calif.: Benjamin Cummings. ISBN .
- ^ Kappock, TJ; Ealick, SE; Stubbe, J (Ekim 2000). "Modular evolution of the purine biosynthetic pathway". Current Opinion in Chemical Biology. 4 (5). s. 567–572. doi:10.1016/s1367-5931(00)00133-2. (PMID) 11006546.
- ^ Sampei, G; Baba, S; Kanagawa, M; Yanai, H; Ishii, T; Kawai, H; Fukai, Y; Ebihara, A; Nakagawa, N; Kawai, G (Ekim 2010). "Crystal structures of glycinamide ribonucleotide synthetase, PurD, from thermophilic eubacteria". Journal of Biochemistry. 148 (4). s. 429–438. doi:10.1093/jb/mvq088. (PMID) 20716513.
- ^ Hoskins, AA; Anand, R; Ealick, SE; Stubbe, J (17 Ağustos 2004). "The formylglycinamide ribonucleotide amidotransferase complex from Bacillus subtilis: metabolite-mediated complex formation". Biochemistry, 32 (43 bas.). s. 10314–27. doi:10.1021/bi049127h. (PMID) 15301530.
- ^ Mueller, EJ; Meyer, E; Rudolph, J; Davisson, VJ; Stubbe, J (1 Mart 1994). "N5-carboxyaminoimidazole ribonucleotide: evidence for a new intermediate and two new enzymatic activities in the de novo purine biosynthetic pathway of Escherichia coli". Biochemistry, 8 (33 bas.). s. 2269–78. doi:10.1021/bi00174a038. (PMID) 8117684.
- ^ Firestine, SM; Poon, SW; Mueller, EJ; Stubbe, J; Davisson, VJ (4 Ekim 1994). "Reactions catalyzed by 5-aminoimidazole ribonucleotide carboxylases from Escherichia coli and Gallus gallus: a case for divergent catalytic mechanisms". Biochemistry, 39 (33 bas.). ss. 11927-34. doi:10.1021/bi00205a031. (PMID) 7918411.
- ^ a b Srere, PA (1987). "Complexes of sequential metabolic enzymes". Annual Review of Biochemistry, 1 (56 bas.). s. 89–124. doi:10.1146/annurev.bi.56.070187.000513. (PMID) 2441660.
- ^ Broach, edited by Jeffrey N. Strathern, Elizabeth W. Jones, James R. (1981). The Molecular biology of the yeast Saccharomyces. Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory. ISBN .
- ^ a b O'Donovan, GA; Neuhard, J (Eylül 1970). "Pyrimidine metabolism in microorganisms". Bacteriological Reviews, 3 (34 bas.). s. 278–343. doi:10.1128/MMBR.34.3.278-343.1970. (PMC) 378357 $2. (PMID) 4919542.
- ^ a b c d Geer, Gerald Karp ; responsible for the revision of chapter 15 Peter van der (2004). Cell and molecular biology : concepts and experiments (4., Wiley International bas.). New York: J. Wiley & Sons. ISBN .
- ^ a b Griffiths, Anthony J. F. (1999). Modern genetic analysis (2. bas.). New York: Freeman. ISBN .
- ^ a b Wu, G (Mayıs 2009). "Amino acids: metabolism, functions, and nutrition". Amino Acids, 1. s. 1–17. doi:10.1007/s00726-009-0269-0. (PMID) 19301095.
- ^ Mousdale, D. M.; Coggins, J. R. (1991). Amino Acid Synthesis. Target Sites for Herbicide Action. s. 29–56. doi:10.1007/978-1-4899-2433-9_2. ISBN .
- ^ Miflin, B. J.; Lea, P. J. (1977). "Amino Acid Metabolism". Annual Review of Plant Physiology, 28. s. 299–329. doi:10.1146/annurev.pp.28.060177.001503.
- ^ a b c Umbarger, HE (1978). "Amino acid biosynthesis and its regulation". Annual Review of Biochemistry. 47 (1). s. 532–606. doi:10.1146/annurev.bi.47.070178.002533. (PMID) 354503.
- ^ Pérez-Arellano, I; Carmona-Alvarez, F; Martínez, AI; Rodríguez-Díaz, J; Cervera, J (Mart 2010). "Pyrroline-5-carboxylate synthase and proline biosynthesis: from osmotolerance to rare metabolic disease". Protein Science. 19 (3). s. 372–82. doi:10.1002/pro.340. (PMC) 2866264 $2. (PMID) 20091669.
- ^ Xu, Y; Labedan, B; Glansdorff, N (Mart 2007). "Surprising arginine biosynthesis: a reappraisal of the enzymology and evolution of the pathway in microorganisms". Microbiology and Molecular Biology Reviews. 71 (1). s. 36–47. doi:10.1128/MMBR.00032-06. (PMC) 1847373 $2. (PMID) 17347518.
- ^ "MetaCyc: L-lysine biosynthesis I". 4 Mayıs 2022 tarihinde kaynağından . Erişim tarihi: 15 Nisan 2022.
- ^ PETERKOFSKY, B; GILVARG, C (Mayıs 1961). "N-Succinyl-L-diaminopimelic-glutamic transaminase". The Journal of Biological Chemistry. 236 (5). s. 1432–8. doi:10.1016/S0021-9258(18)64192-4. (PMID) 13734750.
- ^ KINDLER, SH; GILVARG, C (Aralık 1960). "N-Succinyl-L-2,6-diaminopimelic acid deacylase". The Journal of Biological Chemistry. Cilt 235. s. 3532–3535. doi:10.1016/S0021-9258(18)64502-8. (PMID) 13756049.
- ^ Born, TL; Blanchard, JS (Ekim 1999). "Structure/function studies on enzymes in the diaminopimelate pathway of bacterial cell wall biosynthesis". Current Opinion in Chemical Biology. 3 (5). s. 607–613. doi:10.1016/s1367-5931(99)00016-2. (PMID) 10508663.
- ^ "Escherichia coli K-12 substr. MG1655". serine biosynthesis. SRI International. 15 Ağustos 2016 tarihinde kaynağından . Erişim tarihi: 12 Aralık 2013.
- ^ Bell, JK; Grant, GA; Banaszak, LJ (30 Mart 2004). "Multiconformational states in phosphoglycerate dehydrogenase". Biochemistry. 43 (12). s. 3450–8. doi:10.1021/bi035462e. (PMID) 15035616.
- ^ Dubnovitsky, AP; Kapetaniou, EG; Papageorgiou, AC (Ocak 2005). "Enzyme adaptation to alkaline pH: atomic resolution (1.08 A) structure of phosphoserine aminotransferase from Bacillus alcalophilus". Protein Science. 14 (1). s. 97–110. doi:10.1110/ps.041029805. (PMC) 2253317 $2. (PMID) 15608117.
- ^ Wang, W; Kim, R; Jancarik, J; Yokota, H; Kim, SH (10 Ocak 2001). "Crystal structure of phosphoserine phosphatase from Methanococcus jannaschii, a hyperthermophile, at 1.8 A resolution". Structure. 9 (1). s. 65–71. doi:10.1016/s0969-2126(00)00558-x. (PMID) 11342136.
- ^ Monschau, N; Stahmann, KP; Sahm, H; McNeil, JB; Bognar, AL (1 Mayıs 1997). "Identification of Saccharomyces cerevisiae GLY1 as a threonine aldolase: a key enzyme in glycine biosynthesis". FEMS Microbiology Letters. 150 (1). s. 55–60. doi:10.1111/j.1574-6968.1997.tb10349.x. (PMID) 9163906.
- ^ Pye, VE; Tingey, AP; Robson, RL; Moody, PC (24 Eylül 2004). "The structure and mechanism of serine acetyltransferase from Escherichia coli". The Journal of Biological Chemistry. 279 (39). ss. 40729-36. doi:10.1074/jbc.M403751200. (PMID) 15231846.
- ^ Huang, B; Vetting, MW; Roderick, SL (Mayıs 2005). "The active site of O-acetylserine sulfhydrylase is the anchor point for bienzyme complex formation with serine acetyltransferase". Journal of Bacteriology. 187 (9). s. 3201–5. doi:10.1128/JB.187.9.3201-3205.2005. (PMC) 1082839 $2. (PMID) 15838047.
- ^ McPhalen, CA; Vincent, MG; Picot, D; ; Lesk, AM; Chothia, C (5 Eylül 1992). "Domain closure in mitochondrial aspartate aminotransferase". Journal of Molecular Biology. 227 (1). s. 197–213. doi:10.1016/0022-2836(92)90691-C. (PMID) 1522585.
- ^ Larsen, TM; Boehlein, SK; Schuster, SM; Richards, NG; Thoden, JB; Holden, HM; Rayment, I (7 Aralık 1999). "Three-dimensional structure of Escherichia coli asparagine synthetase B: a short journey from substrate to product". Biochemistry. 38 (49). ss. 16146-57. CiteSeerX 10.1.1.453.5998 $2. doi:10.1021/bi9915768. (PMID) 10587437.
- ^ Velasco, AM; Leguina, JI; Lazcano, A (October 2002). "Molecular evolution of the lysine biosynthetic pathways". Journal of Molecular Evolution. 55 (4). ss. 445-59. Bibcode:2002JMolE..55..445V. doi:10.1007/s00239-002-2340-2. (PMID) 12355264.
- ^ Kotaka, M; Ren, J; Lockyer, M; Hawkins, AR; Stammers, DK (20 Ekim 2006). "Structures of R- and T-state Escherichia coli aspartokinase III. Mechanisms of the allosteric transition and inhibition by lysine". The Journal of Biological Chemistry. 281 (42). ss. 31544-52. doi:10.1074/jbc.M605886200. (PMID) 16905770.
- ^ Hadfield, A; Kryger, G; Ouyang, J; Petsko, GA; Ringe, D; Viola, R (18 Haziran 1999). "Structure of aspartate-beta-semialdehyde dehydrogenase from Escherichia coli, a key enzyme in the aspartate family of amino acid biosynthesis". Journal of Molecular Biology. 289 (4). ss. 991-1002. doi:10.1006/jmbi.1999.2828. (PMID) 10369777.
- ^ Mirwaldt, C; Korndörfer, I; Huber, R (10 Şubat 1995). "The crystal structure of dihydrodipicolinate synthase from Escherichia coli at 2.5 A resolution". Journal of Molecular Biology. 246 (1). ss. 227-39. doi:10.1006/jmbi.1994.0078. (PMID) 7853400.
- ^ Cirilli, M; Zheng, R; Scapin, G; Blanchard, JS (16 Eylül 2003). "The three-dimensional structures of the Mycobacterium tuberculosis dihydrodipicolinate reductase-NADH-2,6-PDC and -NADPH-2,6-PDC complexes. Structural and mutagenic analysis of relaxed nucleotide specificity". Biochemistry. 42 (36). ss. 10644-50. doi:10.1021/bi030044v. (PMID) 12962488.
- ^ Beaman, TW; Binder, DA; Blanchard, JS; Roderick, SL (21 Ocak 1997). "Three-dimensional structure of tetrahydrodipicolinate N-succinyltransferase". Biochemistry. 36 (3). ss. 489-94. doi:10.1021/bi962522q. (PMID) 9012664.
- ^ Weyand, S; Kefala, G; Weiss, MS (30 Mart 2007). "The three-dimensional structure of N-succinyldiaminopimelate aminotransferase from Mycobacterium tuberculosis". Journal of Molecular Biology. 367 (3). ss. 825-38. doi:10.1016/j.jmb.2007.01.023. (PMID) 17292400.
- ^ Nocek, BP; Gillner, DM; Fan, Y; Holz, RC; Joachimiak, A (2 Nisan 2010). "Structural basis for catalysis by the mono- and dimetalated forms of the dapE-encoded N-succinyl-L,L-diaminopimelic acid desuccinylase". Journal of Molecular Biology. 397 (3). ss. 617-26. doi:10.1016/j.jmb.2010.01.062. (PMC) 2885003 $2. (PMID) 20138056.
- ^ Pillai, B; Cherney, M; Diaper, CM; Sutherland, A; Blanchard, JS; Vederas, JC; James, MN (23 Kasım 2007). "Dynamics of catalysis revealed from the crystal structures of mutants of diaminopimelate epimerase". Biochemical and Biophysical Research Communications. 363 (3). ss. 547-53. doi:10.1016/j.bbrc.2007.09.012. (PMID) 17889830.
- ^ Gokulan, K; Rupp, B; Pavelka MS, Jr; Jacobs WR, Jr; Sacchettini, JC (16 Mayıs 2003). "Crystal structure of Mycobacterium tuberculosis diaminopimelate decarboxylase, an essential enzyme in bacterial lysine biosynthesis". The Journal of Biological Chemistry. 278 (20). ss. 18588-96. doi:10.1074/jbc.M301549200. (PMID) 12637582.
- ^ a b c Weaver, Robert F. (2005). Molecular biology (3. bas.). Boston: McGraw-Hill Higher Education. ISBN .
- ^ a b c d e Cooper, Geoffrey M. (2000). The Cell: A Molecular Approach (2. bas.). Washington (DC): ASM Press. ISBN .
- ^ Jackson, R.J. (Şubat 2010). "The mechanism of eukaryotic translation initiation and principles of its regulation". Molecular Cell Biology. 10 (2). ss. 113-127. doi:10.1038/nrm2838. (PMC) 4461372 $2. (PMID) 20094052.
- ^ Green, Rachel; Harry F. Noller (1997). "Ribosomes and Translation". Annu. Rev. Biochem. Cilt 66. ss. 679-716. doi:10.1146/annurev.biochem.66.1.679. (PMID) 9242921.
- ^ a b c d Weissbach, Herbert; Pestka, Sidney (1977). Molecular Mechanisms of Protein Biosynthesis. New York: Academic Press. ISBN .
- ^ Frank, J; Haixiao Gao (Eylül 2007). "The process of mRNA–tRNA translocation". PNAS. 104 (50). ss. 19671-19678. doi:10.1073/pnas.0708517104. (PMC) 2148355 $2. (PMID) 18003906.
- ^ a b c Bandeali, Salman J.; Daye, Jad; Virani, Salim S. (30 Kasım 2013). "Novel Therapies for Treating Familial Hypercholesterolemia". Current Atherosclerosis Reports. 16 (1). s. 382. doi:10.1007/s11883-013-0382-0. (PMID) 24293346.
- ^ a b c Kang, Tae Hyuk; Park, Yongjin; Bader, Joel S.; Friedmann, Theodore; Cooney, Austin John (9 Ekim 2013). "The Housekeeping Gene Hypoxanthine Guanine Phosphoribosyltransferase (HPRT) Regulates Multiple Developmental and Metabolic Pathways of Murine Embryonic Stem Cell Neuronal Differentiation". PLOS ONE. 8 (10). ss. e74967. Bibcode:2013PLoSO...874967K. doi:10.1371/journal.pone.0074967. (PMC) 3794013 $2. (PMID) 24130677.
- ^ a b c Walport, Ken Murphy, Paul Travers, Mark (2011). Janeway's Immunobiology. 8. Oxford: Taylor & Francis. ISBN .
- ^ a b Hughes, edited by Donald C. Lo, Robert E. (2010). Neurobiology of Huntington's disease : applications to drug discovery. 2nd. Boca Raton: CRC Press/Taylor & Francis Group. ISBN .
- ^ Biglan, Kevin M.; Ross, Christopher A.; Langbehn, Douglas R.; Aylward, Elizabeth H.; Stout, Julie C.; Queller, Sarah; Carlozzi, Noelle E.; Duff, Kevin; Beglinger, Leigh J.; Paulsen, Jane S. (26 Haziran 2009). "Motor abnormalities in premanifest persons with Huntington's disease: The PREDICT-HD study". Movement Disorders. 24 (12). ss. 1763-1772. doi:10.1002/mds.22601. (PMC) 3048804 $2. (PMID) 19562761.
wikipedia, wiki, viki, vikipedia, oku, kitap, kütüphane, kütübhane, ara, ara bul, bul, herşey, ne arasanız burada,hikayeler, makale, kitaplar, öğren, wiki, bilgi, tarih, yukle, izle, telefon için, turk, türk, türkçe, turkce, nasıl yapılır, ne demek, nasıl, yapmak, yapılır, indir, ücretsiz, ücretsiz indir, bedava, bedava indir, mp3, video, mp4, 3gp, jpg, jpeg, gif, png, resim, müzik, şarkı, film, film, oyun, oyunlar, mobil, cep telefonu, telefon, android, ios, apple, samsung, iphone, xiomi, xiaomi, redmi, honor, oppo, nokia, sonya, mi, pc, web, computer, bilgisayar
Biyosentez substratlarin canli organizmalarda daha karmasik urunlere donusturuldugu cok asamali enzim katalizli bir surectir Biyosentezde basit bilesikler modifiye edilir diger bilesiklere donusturulur veya makromolekuller olusturmak uzere birlestirilir Bu surec genellikle metabolik yollardan olusur Bu biyosentetik yollardan bazilari tek bir hucresel organel icinde yer alirken digerleri birden fazla hucresel organel icinde yer alan enzimleri icerir Bu biyosentetik yollarin ornekleri arasinda cift katli lipit katmaninin bilesenlerinin ve nukleotidlerin uretimi yer alir Biyosentez genellikle anabolizma ile es anlamlidir ve bazi durumlarda birbirinin yerine kullanilir Biyosentez olaylarinin gerceklesmesi icin oncu bilesikler kimyasal enerji ornegin ATP ve koenzime sahip katalitik enzimler gereklidir Bunlar makromolekullerin yapi taslari olan monomerleri olusturur Bazi onemli makromolekuller peptit baglari yoluyla birlestirilen ve amino asit monomerlerinden olusan proteinleri ve fosfodiester baglari yoluyla birlestirilen nukleotitlerden olusan DNA molekullerini icermektedir Kimyasal reaksiyonlarin ozellikleriBiyosentez bir dizi kimyasal reaksiyonlar ile gerceklesir Bu reaksiyonlarin gerceklesmesi icin asagidaki ozellikler ve unsurlar gereklidir Bu bilesikler bir reaksiyondaki baslangic molekulleri veya substratlaridir Bunlar ayrica belirli bir kimyasal islemde reaktanlar olarak da gorev yapabilirler Kimyasal enerji Kimyasal enerji yuksek enerjili molekuller seklinde bulunabilir Bu molekuller enerjik olarak elverissiz reaksiyonlar icin gereklidir Bu bilesiklerin hidrolizi bir reaksiyonu ileriye tasir ATP gibi yuksek enerjili molekuller uc fosfata sahiptir Cogu zaman terminal fosfat hidroliz sirasinda ATP den ayrilir ve baska bir molekule aktarilir Katalitik enzimler Bu molekuller reaksiyonun hizini artirarak ve aktivasyon enerjisini dusurerek bir reaksiyonu katalize eden ozel proteinlerdir Kofaktor Koenzim Kofaktorler kimyasal reaksiyonlara yardimci olan molekullerdir Bunlar metal iyonlari NADH ve asetil CoA gibi vitamin turevleri veya ATP gibi vitamin olmayan turevler olabilir En basit anlamiyla biyosentezde meydana gelen reaksiyonlar su formattadir Reaktant enzimU ru n displaystyle ce Reaktant gt enzim ddot U r ddot u n dd Bu temel denklemin ayrintili olarak tartisilabilir bazi varyasyonlari sunlardir Genellikle cok asamali bir reaksiyon yolunun parcasi olarak diger bilesiklere donusturulen basit bilesikler Bu tip reaksiyonun iki ornegi nukleik asitlerin olusumunda ve translasyondan once tRNA nin yuklenmesi sirasinda meydana gelir Bu adimlardan bazilari icin kimyasal enerji gereklidir O ncu moleku l ATP AMP PPi displaystyle ce ddot O nc ddot u molek ddot u l ATP lt gt AMP PP i dd Kofaktorlerin yardimiyla baska bilesiklere donusturulen basit bilesikler Ornegin fosfolipitlerin sentezi asetil CoA yi gerektirirken baska bir zar bileseni olan sfingolipidlerin sentezi sfingozin omurgasinin olusumu icin NADH ve FADH yi gerektirir Bu ornekler icin genel denklem su sekildedir O ncu moleku l Kofakto r enzimmakromoleku l displaystyle ce ddot O nc ddot u molek ddot u l Kofakt ddot o r gt enzim makromolek ddot u l dd Bir makromolekul olusturmak icin birlesen basit bilesikler Ornegin yag asitleri birleserek fosfolipitleri olusturur Buna karsilik fosfolipidler ve kolesterol lipid cift tabakasini olusturmak icin kovalent olmayan baglarla etkilesime girer Bu reaksiyon asagidaki gibi gosterilebilir moleku l 1 moleku l 2 makromoleku l displaystyle ce molek ddot u l 1 molek ddot u l 2 gt makromolek ddot u l dd LipitLipit membran tabakasi Bircok karmasik makromolekul basit tekrarlanan yapilarin bir modelinde sentezlenir Ornegin lipitlerin en basit yapilari yag asitleridir Yag asitleri hidrokarbon turevleridir bir karboksil grubu bas ve bir hidrokarbon zinciri kuyruk icerir Bu yag asitleri daha buyuk bilesenler olusturur ve bunlar da lipit cift tabakasini olusturmak icin kovalent olmayan etkilesimleri icerir Yag asidi zincirleri membran lipidlerinin iki ana bileseni olan fosfolipitler ve sfingolipidlerde bulunur Ucuncu bir ana zar bileseni olan kolesterol bu yag asidi birimlerini icermez Fosfolipitler Tum biyomembranlarin temeli iki katmanli bir fosfolipit yapisindan olusur Fosfolipid molekulu amfifildir hidrofilik bir kutup basi ve hidrofobik kutupsuz bir kuyruk icerir Fosfolipid kafalari birbirleriyle ve sulu ortamla etkilesime girerken hidrokarbon kuyruklari kendilerini merkezde sudan uzaga yonlendirir Bu son etkilesimler iyonlar ve molekuller icin bir bariyer gorevi goren cift katmanli yapiyi yonlendirir Cesitli fosfolipid turleri vardir ancak sentez yollari farklidir Fosfolipid sentezinde ilk adim endoplazmik retikulumda ve mitokondride gerceklesir Sentez sirasinda veya da olusur Sentez su sekilde gerceklesmektedir Sentez tarafindan saglanan bir yag asidi zincirinin eklenmesi yoluyla lizofosfatidata donusturulen gliserol 3 fosfat ile baslar Daha sonra lizofosfatidat ikinci bir acil CoA nin katkida bulundugu baska bir yag asidi zincirinin eklenmesi yoluyla fosfatidata donusturulur bu adimlarin tumu gliserol fosfat enzimi tarafindan katalize edilir Fosfolipid sentezi endoplazmik retikulumda devam eder ve biyosentez belirli fosfolipidin bilesenlerine bagli olarak farklilik gosterir Sfingolipitler Fosfolipidler gibi bu yag asidi turevlerinin de bir kutup basi ve kutupsal olmayan kuyruklari vardir Fosfolipidlerin aksine sfingolipidlerin bir sfingozin omurgasi vardir Sfingolipidler okaryotik hucrelerde ozellikle merkezi sinir sisteminde bol miktarda bulunur Ornegin sfingomyelin sinir lifleri miyelin kilifinin bir parcasidir Sfingolipidler bir sfingozin omurgasinin amino grubuna bagli bir yag asidi zincirinden olusan seramidlerden olusur Bu seramidler sfingosinin asilasyonundan sentezlenir Sfingosinin biyosentezi su sekildedir Sfingozin sentezi sirasinda ve serin dehidrosfingosin olusumuyla sonuclanan bir kondenzasyon reaksiyonuna girer Bu urun daha sonra FAD tarafindan oksidasyon reaksiyonu yoluyla sfingosine donusturulen dihidrospingozini olusturmak uzere indirgenir Kolesterol Kolesterol sterol adi verilen bir molekul sinifina aittir Sterollerin dort kaynasmis halkasi ve bir hidroksil grubu vardir Kolesterol ozellikle onemli bir molekuldur Sadece lipid zarlarinin bir bileseni olarak gorev yapmakla kalmaz ayni zamanda kortizol testosteron ve ostrojen de dahil olmak uzere cesitli steroid hormonlarinin yapisina katilir Kolesterol asetil CoA dan sentezlenir Sentezi su sekildedir Daha genel olarak bu sentez uc asamada gerceklesir Birinci asama sitoplazmada gerceklesir ve ikinci ve ucuncu asama endoplazmik retikulumda meydana gelir Asamalari ise su sekildedir Kolesterolun yapi tasi olan sentezi Alti molekul izopentenil fosfatin yogunlasmasiyla skualen olusumu Birkac enzimatik reaksiyon yoluyla skualenin kolesterole donusumuNukleotitNukleotitlerin biyosentezi substratlari daha karmasik urunlere donusturen enzim katalizli reaksiyonlardan olusmaktadir Nukleotitler DNA ve RNA nin yapi taslaridir Nukleotitler RNA daki riboz sekerinden ve DNA daki deoksiriboz sekerden olusan bes uyeli bir halkadan olusur bu sekerler bir glikosidik bag ve bir fosfat grubu ile bir purin veya pirimidin bazina baglanir Purinler IMP sentezi DNA nukleotidlerinden adenozin ve bir glikosidik bag ile bir riboz sekerine bagli bir purin bazindan olusur RNA nukleotidleri deoksiadenozin ve deoksiguanozin durumunda purin bazlari bir glikozidik bag ile bir deoksiriboz sekere baglanir DNA ve RNA nukleotidlerindeki purin bazlari cogu tek hucreli organizmada bulunan on iki asamali bir reaksiyon mekanizmasinda sentezlenir Daha gelismis okaryotlar on reaksiyon adiminda benzer bir reaksiyon mekanizmasi kullanir Purin bazlari fosforibosil pirofosfatin PRPP purin baz biyosentezindeki ilk anahtar ara madde olan inozin monofosfata IMP donusturulmesiyle sentezlenir IMP nin daha fazla enzimatik modifikasyonu nukleotitlerin adenozin ve guanozin bazlarini uretir Purinlerin biyosentezi su sekilde gerceklesir Purin biyosentezindeki ilk adim tarafindan gerceklestirilen bir yogunlasma reaksiyonudur Bu enzim amino grubunu glutaminden PRPP ye aktararak olusturur Asagidaki adim ATP den bir fosfat grubunun eklenmesiyle glisinin aktivasyonunu gerektirir GAR sentetaz aktiflestirilmis glisinin PRPP uzerinde yogunlastirilmasini gerceklestirerek glisinamid ribonukleotid GAR olusturur GAR transformilaz GAR in amino grubuna bir formil grubu ekleyerek formilglisinamid ribonukleotid FGAR olusturur FGAR amidotransferaz bir nitrojen grubunun FGAR a eklenmesini katalize ederek formilglisinamidin ribonukleotid FGAM olusturur FGAM siklaz 5 li halkasini 5 aminoimidazol ribonukleotidi AIR olusturan bir su molekulunun cikarilmasini iceren halka kapanmasini katalize eder N5 CAIR sentetaz bir karboksil grubunu transfer ederek ara urun olan N5 karboksiaminoimidazol ribonukleotidi N5 CAIR olusturur N5 CAIR mutazi karboksil fonksiyonel grubunu yeniden duzenler ve onu imidazol halkasina aktararak karboksiamino imidazol ribonukleotidi CAIR olusturur AIR den CAIR olusumunun iki asamali mekanizmasi cogunlukla tek hucreli organizmalarda bulunur Daha yuksek okaryotlar bir karboksil grubunu dogrudan AIR imidazol halkasina aktaran ve CAIR olusturan AIR karboksilaz enzimini icerir SAICAR sentetaz aspartat ile imidazol halkasinin eklenen karboksil grubu arasinda SAICAR olusturan bir peptit bagi olusturur SAICAR liyaz eklenen aspartatin karbon iskeletini cikararak amino grubunu terk eder ve AICAR olusturur AICAR transformilaz bir karbonil grubunu AICAR a aktararak FAICAR olusturur Son adim purin halkasinin kapanmasini gerceklestiren ve inozin monofosfat ara urununu olusturan IMP sentaz enzimi ile gerceklesir Pirimidinler Uridin monofosfat UMP biyosentezi Bir glikozidik bag yoluyla riboz sekerine baglanan diger DNA ve RNA nukleotid bazlari timin sitozin ve urasildir Urasil RNA ya ozgudur UMP biyosentezinde mitokondriyal ic zarda bulunan bir enzim ve sitozolde bulunan cok islevli enzimler bulunmaktadir Pirimdinlerin biyosentezi su sekilde gerceklesir Ilk adimda karbamoil fosfat olusturmak icin ATP ye bagli bir reaksiyonda glutamin ile CO2 yi birlestiren enzim bulunmaktadir uridosuksinat olusturmak icin karbamoil fosfati aspartat ile yogunlastirir olusturmak icin su kaybeden bir reaksiyon olan halka kapanmasini gerceklestirir Mitokondriyal ic zar icinde yer alan dihidroorotati oksitler Orotat fosforibosil hidrolaz OMP pirofosforilaz orotati PRPP ile yogunlastirarak orotidin 5 fosfat olusturur orotidin 5 fosfatin UMP ye donusumunu katalize eder Uridin nukleotid bazi sentezlendikten sonra diger bazlar olan sitozin ve timin sentezlenir Sitozin biyosentezi UMP nin ye donusumunu iceren iki asamali bir reaksiyondur UMP ye fosfat ilavesi bir kinaz enzimi tarafindan katalize edilir enzimi bir sonraki reaksiyon adimini katalize eder bir amino grubunu glutaminden uridine transfer ederek UTP nin ye donusturulmesi CTP nin sitozin bazini olusturur UTP ATP glutamin CTP ADP glutamat reaksiyonu su sekildedir Sitozin hem DNA hem de RNA da bulunan bir nukleotittir Ancak urasil sadece RNA da bulunur Bu nedenle UTP sentezlendikten sonra DNA ya dahil edilmesi icin formuna donusturulmesi gerekir Bu donusum enzimi ile gerceklesir Deoksiriboz olusturmak icin riboz sekerinin 2 OH sini uzaklastiran bu reaksiyon sekere bagli bazlardan etkilenmez Bu ozgul olmama ribonukleozit trifosfat reduktazin benzer bir mekanizma ile tum nukleotid trifosfatlari deoksiribonukleotite donusturmesine izin verir Urasilin aksine timin bazlari cogunlukla RNA da degil DNA da bulunur Hucreler normalde RNA daki riboz sekerlerine bagli timin bazlari icermez bu da hucrelerin yalnizca deoksiriboza bagli timini sentezledigini gosterir Timidilat sentetaz enzimi timin kalintilarinin dUMP den dTMP ye sentezlenmesinden sorumludur Bu reaksiyon dTMP olusturmak icin bir metil grubunu dUMP nin urasil bazina aktarir Timidilat sentaz reaksiyonu su sekildedir dUMP 5 10 metilentetrahidrofolat dTMP dihidrofolatDNADNA polimeraz sablon iplik boyunca 3 ila 5 yonunde hareket ederken 5 ila 3 yonunde yeni bir iplik sentezler Okaryotik ve prokaryotik canlilarda DNA sentezi arasinda farkliliklar olmasina ragmen asagidaki bolumler her iki organizma tarafindan paylasilan DNA replikasyonunun temel ozelliklerini anlatmaktadir DNA fosfodiester baglari ile birlestirilen nukleotitlerden olusur Cekirdekte gerceklesen DNA sentezi yari koruyucu bir surectir Bu elde edilen DNA molekulunun ana yapidan orijinal bir iplik ve yeni bir iplik icerdigi anlamina gelir DNA sentezi dort deoksinukleosit trifosfat bir sablon iplik ve nukleotitlerin dahil edilecegi serbest 3 OH li bir primer gerektiren bir DNA polimeraz ailesi tarafindan katalize edilir DNA replikasyonunun gerceklesmesi icin DNA sarmalini cozen helikaz adi verilen enzimler tarafindan bir replikasyon catali olusturulur Replikasyon catalindaki topoizomerazlar DNA nin cozulmesinin neden oldugu super bobinleri kaldirir ve tek iplikli DNA baglayici proteinler replikasyondan once stabilize edilmis iki tek iplikli DNA sablonunu korur DNA sentezi serbest bir 3 OH ile bir RNA primeri yapan RNA polimeraz primeri tarafindan baslatilir Bu primer tek iplikli DNA sablonuna baglidir ve DNA polimeraz nukleotidleri dahil ederek zinciri uzatir DNA polimeraz ayrica yeni sentezlenen DNA zincirini de duzeltir DNA polimeraz tarafindan katalize edilen polimerizasyon reaksiyonu sirasinda buyuyen zincirin 3 OH si tarafindan bir deoksinukleosit trifosfatin en icteki fosfor atomu uzerinde bir nukleofilik saldiri meydana gelir bu yeni bir nukleotidi baglayan ve pirofosfat salan bir fosfodiester baginin olusumunu saglar Replikasyon sirasinda ayni anda surekli sentezlenen ve replikasyon catalina dogru buyuyen oncu iplik ve Okazaki parcalarinda sureksiz olarak yapilan ve replikasyon catalindan uzaklasan gecikmeli iplik olmak uzere iki tip iplik olusur Okazaki parcalari surekli bir iplik olusturmak icin DNA ligaz ile ve kovalent bag ile birlestirilir Daha sonra DNA replikasyonunu tamamlamak icin RNA primerleri cikarilir ve olusan bosluklar DNA ile degistirilir ve DNA ligaz yoluyla birlestirilir Amino asitProtein peptid baglariyla birbirine baglanan amino asitlerden olusan bir polimerdir Dogada bulunan 300 den fazla amino asit vardir Bunlardan sadece yirmi tanesi standart temel amino asitler olarak bilinir ve proteinin yapi taslaridir Sadece yesil bitkiler ve cogu mikrop tum canli turlerinin ihtiyac duydugu 20 standart amino asidin tamamini sentezleyebilir Memeliler yirmi standart amino asitten yalnizca onunu sentezleyebilir Diger amino asitler olan valin metionin losin izolosin fenilalanin lizin treonin ve triptofan histidin ve arginin beslenme yoluyla elde edilir Amino asidin temel yapisi L amino asit Standart amino asitlerin genel yapisi bir birincil amino grubu bir karboksil grubu ve bagli fonksiyonel grubu icerir Amino asitler fonksiyonel gruplar nedeniyle birbirinden ayrilir a karbona bagli uc farkli grubun bir sonucu olarak amino asitler asimetrik molekullerdir Glisin haric tum standart amino asitler icin a karbon bir kiral merkezdir Glisin durumunda a karbon iki hidrojen atomuna sahiptir bu durum molekule simetri ekler Prolin haric yasamda bulunan tum amino asitler L izoform konformasyonuna sahiptir Prolin amino grubu ile bir halka olusturan a karbon uzerinde fonksiyonel bir gruba sahiptir Glutamin oksoglutarat aminotransferaz ve glutamin sentetazAmino asitlerin azot kaynagi Amino asit biyosentezindeki onemli bir adim a karbon uzerine bir azot grubunun dahil edilmesidir Hucrelerde azot gruplarini dahil etmenin iki ana yolu vardir Birinci yol glutaminin amit amino grubunu uzaklastiran ve onu uzerine transfer eden ve iki glutamat molekulu ureten GOGAT enzimi ile dahil etmektir Bu kataliz reaksiyonunda azot kaynagi olarak glutamin gorev yapar Bu reaksiyon yukaridaki resimde gosterilmistir Amino asitlerin a karbonuna azot dahil etmenin diger yolu GDH enzimi ile dahil etmektir GDH amonyagi 2 oksoglutarat uzerine aktarabilir ve glutamat olusturabilir Ayrica GS enzimi amonyagi glutamata aktarabilir ve glutamini yenileyerek glutamin sentezleyebilir Amino asitlerin glutamat ailesi Amino asitlerin ailesi amino asit glutamattan turetilen amino asitleri icerir Bu aile glutamat glutamin prolin ve argininden olusmaktadir Bu aile ayrica turetilen amino asit lizini de icerir Glutamat ve glutamin biyosentezi azot asimilasyonunda onemli bir adimdir GOGAT ve GDH enzimleri azot asimilasyon reaksiyonlarini katalize eder Glutamat ve glutamin biyosentezi azot asimilasyonunda onemli bir adimdir GOGAT ve GDH enzimleri azot asimilasyon reaksiyonlarini katalize eder Bakterilerde enzimi bir fosfat grubunu ATP den glutamata aktararak prolin biyosentezini baslatir Sonraki reaksiyon L glutamat 5 fosfatin grubunun indirgenmesini katalize eden enzim P5CS tarafindan katalize edilir Bu kendiliginden pirolin 5 karboksilata siklize olan glutamat semialdehit olusumu ile sonuclanir Pirolin 5 karboksilat bir prolin amino asidi verecek sekilde pirolin 5 karboksilat reduktaz P5CR enzimi tarafindan daha da indirgenir Bakterilerde arginin biyosentezinin ilk adiminda asetil grubunun N a pozisyonunda asetil CoA dan aktarilmasiyla glutamat asetillenir bu kendiliginden siklizasyonu onler N asetilglutamat sentaz enzimi glutamat N asetiltransferaz asetilasyon adimini katalize etmekten sorumludur Sonraki adimlar ve asetilornitin suksinildiamino pimelat aminotransferaz enzimleri tarafindan katalize edilir ve N asetil L ornitin verir Asetillornitin asetil grubu AO veya OAT enzimi tarafindan cikarilir ve bu verir Daha sonra ve enzimleri ornitini arginine donusturur Diaminopimelik asit yolu Iki farkli lizin biyosentetik yolu vardir Diaminopimelik asit yolu ve a aminoadipat yolu olmak uzere ikiye ayrilir Iki sentetik yoldan en yaygin olani lizin elde etmek icin aspartata karbon gruplari ekleyen birkac enzimatik reaksiyondan olusur Aspartat kinaz aspartati fosforile ederek ve aspartil fosfat ureterek diaminopimelik asit yolunu baslatir Aspartat semialdehit dehidrojenaz aspartil fosfatin NADPH ye bagli indirgenmesini katalize ederek aspartat semialdehit verir 4 hidroksi tetrahidrodipikolinat sentaz b aspartil 4 semialdehite bir piruvat grubu ekler ve bir su molekulu cikarilir Bu neden olur ve 2S 4S 4 hidroksi 2 3 4 5 tetrahidrodipikolinata yol acar 4 hidroksi tetrahidrodipikolinat reduktaz 2S 4S 4 hidroksi 2 3 4 5 tetrahidrodipikolinatin NADPH tarafindan indirgenmesini katalize ederek D piperidein 2 6 dikarboksilat 2 3 4 5 tetrahidrodipikolinat ve H20 ortaya cikarir Tetrahidrodipikolinat asiltransferaz halka acilmasiyla sonuclanan asetilasyon reaksiyonunu katalize eder ve N asetil a amino e ketopimelat verir N asetil a amino e ketopimelat in keto grubunu uzaklastiran ve N suksinil L diaminopimelat verecek sekilde bir amino grubuyla degistiren transaminasyon reaksiyonunu katalize eder N asildiaminopimelat deasilaz L L diaminopimelat verecek sekilde N suksinil L diaminopimelatin deasilasyonunu katalize eder L L diaminopimelatin L L diaminopimelatin mezo formuna donusumunu katalize eder karboksil grubunun cikarilmasini katalize ederek L lisin verir Amino asitlerin serin ailesi Amino asidin serin ailesi serin sistein ve glisinden olusmaktadir Cogu mikroorganizma ve bitki amino asit sisteininden metiyonini sentezlemek icin kukurt elde eder Ayrica serinin glisine donusumu metiyonin ve histidinin biyosentezi icin gerekli karbonlari saglar Serin biyosentezi sirasinda fosfogliserat dehidrojenaz enzimi 3 fosfo D gliserati oksitleyerek veren ilk reaksiyonu katalize eder Reaksiyonda bir amino grubunu glutamattan 3 fosfonooksipiruvat uzerine transfer etirilerek L fosfoserin veren enzim tarafindan katalize edilir Son adimda L serin verecek sekilde L fosfoserin i fosforile eden enzim tarafindan katalize edilir Glisin biyosentezi icin bilinen iki yol vardir Bir yolunda ana karbon kaynagi olarak etanol ve asetat kullanan organizmalar glisin sentezlemek icin yolu kullanir Glisin biyosentezinin diger yolu glikolitik yol olarak bilinir Bu yol glikolizin ara urunlerinden sentezlenen serini glisine donusturur Glikolitik yolda enzimi glisin elde etmek icin serinin bolunmesini katalize eder ve bolunmus serin karbon grubunu aktararak olusturur Sistein biyosentezi inorganik kukurtun dahil oldugu iki asamali bir reaksiyondur Mikroorganizmalarda ve bitkilerde enzimi asetil grubunun asetil CoA dan L serine transferini katalize ederek O asetil L serin verir enzimi tarafindan katalize edilen reaksiyon adimi sistein verecek sekilde O asetil L serinin asetil grubunu sulfur ile degistirir Aspartat amino asit ailesi Aspartat amino asit ailesi treonin lizin metionin izolosin ve aspartattan olusmaktadir Lizin ve izolosin karbon iskeletlerinin bir kismi piruvattan turetilmis olsa da aspartat ailesinin bir parcasi olarak kabul edilir Metiyonin durumunda metil karbon serin ve kukurt grubundan turetilir ancak cogu organizmada sisteinden turetilir Aspartat biyosentezi tek bir enzim tarafindan katalize edilen tek adimli bir reaksiyondur Aspartat aminotransferaz enzimi bir amino grubunun aspartattan a ketoglutarat uzerine transferini katalize ederek glutamat ve oksaloasetat verir Asparagin aspartat uzerine bir amino grubunun ATP ye bagimli eklenmesiyle sentezlenir asparagin sentetaz asparagin verecek sekilde glutamin veya cozunur amonyaktan aspartata azot eklenmesini katalize eder Diaminopimelik asit lizin biyosentetik yolu Lizinin diaminopimelik asit biyosentetik yolu aspartat amino asit ailesine aittir Bu yol aspartati lizine donusturen dokuz enzim katalizli reaksiyonu icerir Aspartat kinaz ATP den aspartil b fosfat veren karboksilat grubuna ATP den bir fosfor aktararak diaminopimelik asit yolundaki ilk adimi katalize eder Aspartat semialdehit dehidrojenaz aspartat b semialdehit verecek sekilde aspartil b fosfatin fosforilasyonu yoluyla indirgeme reaksiyonunu katalize eder Dihidrodipikolinat sentaz aspartat b semialdehitin piruvat ile katalize ederek dihidrodipikolinik asit verir dihidrodipikolinik asidin indirgenmesini katalize ederek tetrahidrodipikolinik asit verir bir suksinil grubunun suksinil CoA dan tetrahidrodipikolinik asit uzerine transferini katalize ederek N suksinil L 2 6 diaminoheptandioat verir N suksinildiaminopimelat aminotransferaz bir amino grubunun glutamattan N suksinil L 2 6 diaminoheptandioat uzerine transferini katalize ederek N suksinil L L diaminopimelik asit verir asil grubunun N suksinil L L diaminopimelik asitten cikarilmasini katalize ederek L L diaminopimelik asit verir verecek sekilde L L diaminopimelik asidin a karbonunun ters cevrilmesini katalize eder Siaminopimelat dekarboksilaz karbon dioksit grubunu mezo diaminopimelik asitten uzaklastirarak L lizin veren lizin biyosentezindeki son adimi katalize eder ProteinlerRNA antikodonu bir amino asidi buyuyen polipeptit zincirine baglamak icin mRNA kodonu ile etkilesime girer tRNA sarj islemi Protein sentezi translasyon adi verilen bir surecle gerceklesir Translasyon sirasinda mRNA adi verilen genetik materyal bir protein polipeptit zinciri olusturmak icin ribozomlar tarafindan okunur Bu islem bir ucunda amino asitleri baglayarak ve diger ucunda mRNA ile etkilesime girerek bir adaptor gorevi goren transfer RNA sini tRNA gerektirir tRNA ve mRNA arasindaki ikinci eslesme zincire dogru amino asidin eklenmesini saglar Protein sentezi baslama uzama ve sonlandirma olmak uzere uc asamada gerceklesir Prokaryotik canlilarda translasyon farklidir Translasyon oncesi Translasyon baslamadan once belirli bir amino asidi karsilik gelen tRNA ya baglama islemi gerceklesmelidir tRNA yuklemesi olarak adlandirilan bu reaksiyon tarafindan katalize edilir Belirli bir amino asidin taninmasindan ve yuklenmesinden belirli bir tRNA sentetaz sorumludur Ayrica bu enzim tRNA ile ayni kokenli amino asidi arasinda dogru baglanmayi saglamak icin ozel ayirici bolgelere sahiptir Bir amino asidi karsilik gelen tRNA ya baglamanin ilk adimi aminoasil AMP nin olusumudur Aminoasit ATP Aminoasil AMP PPi displaystyle ce Amino asit ATP lt gt Aminoasil AMP PP i Bunu aminoasil grubunun aminoasil AMP den bir tRNA molekulune transferi izler Ortaya cikan molekul aminoasil tRNA dir Aminoasil AMP tRNA Aminoasil tRNA AMP displaystyle ce Aminoasil AMP tRNA lt gt Aminoasil tRNA AMP Aminoasil tRNA sentetaz tarafindan katalize edilen bu iki adimin kombinasyonu buyuyen polipeptit zincirine amino asitler eklemeye hazir olan yuklu bir tRNA uretir Bir amino asidi baglamaya ek olarak tRNA mRNA da da bulunan kodon adi verilen spesifik nukleotit ucluleri ile baz ciftleri olusturan antikodon adi verilen uc nukleotit birimine sahiptir kodonlar belirli bir amino asidi kodlar Bu etkilesim protein sentezi icin bolge gorevi goren ribozom sayesinde mumkundur Ribozom aminoasil bolgesi A bolgesi peptidil bolgesi P bolgesi ve cikis bolgesi E bolgesi olmak uzere uc tRNA baglanma bolgesine sahiptir Bir mRNA transkriptinde cok sayida kodon vardir Bu nedenle bir amino asidin birden fazla kodon ile etkilesimi cok yaygindir buna dejenerasyon denir Toplamda 20 amino asitten biri icin her kodun 61 i olmak uzere 64 kodon vardir geri kalan kodonlar ise zincir sonlandirmasini belirtir Translasyonun asamalari Translasyonun baslatma uzama ve sonlandirma olmak uzere uc asamasi vardir Translasyon1 Adim Baslatma Baslatma asamasinin tamamlanmasi icin bu uc olayin gerceklesmesi gerekmektedir Ribozomun mRNA ya alinmasi Yuklu bir baslatici tRNA nin ribozomun P bolgesine baglanmasi Ribozomun mRNA nin baslangic kodonu ile dogru hizalanmasi2 Adim Uzama Baslatmayi takiben polipeptit zinciri antikodon kodon etkilesimleri yoluyla uzatilir ve ribozom polipeptit zincirine birer birer amino asitler ekler Amino asitlerin dogru eklenmesini saglamak icin asagidaki adimlar gerceklesmelidir Dogru tRNA nin ribozomun A bolgesine baglanmasi A bolgesindeki tRNA ile P bolgesindeki tRNA ya bagli polipeptit zinciri arasinda peptit baginin olusmasi Uc nukleotid tarafindan tRNA mRNA kompleksinin translokasyonu veya ilerlemesi Translokasyon E bolgesindeki tRNA nin hareketini baslatir ve tRNA yi A bolgesinden P bolgesine kaydirir A bolgesini gelen bir tRNA nin baska bir amino asit eklemesi icin serbest birakir 3 Adim Sonlandirma Translasyonun son asamasi bir durdurma kodonu A bolgesine girdiginde gerceklesir Ardindan asagidaki adimlar gerceklesir P bolgesinde bulunan tRNA dan polipeptit zincirinin hidrolizine neden olan salim faktorleri tarafindan kodonlarin taninmasi Polipeptit zincirinin salinimi Gelecekte gerceklesecek translasyon surecleri icin ribozomun ayrilmasi ve geri donusumu Translasyonda gorev alan onemli molekuller Molekul Bulundugu adim GorevitRNA sentetaz Baslatmadan once tRNA sarjindan sorumludur mRNA Baslatma uzama sonlandirma Protein sentezi icin sablon ve amino asitleri kodlayan kodonlar olarak adlandirilan bolgeleri bulundurur tRNA A P E ribozom bolgelerine baglanir dogru amino asidin buyuyen polipeptit zincirine dahil edilmesini saglamak icin mRNA kodonu ile antikodon baz ciftlerini barindirir Ribozom Protein sentezini yonetir ve peptit bagi olusumunu katalize eder Makromolekul eksikligi ile iiskili hastaliklarAilesel hiperkolesterolemi kolesterol birikimine neden olur Biyosentetik yollardaki hatalar makromolekullerin malformasyonu veya fonksiyonel molekullerin yetersiz uretimi dahil olmak uzere zararli sonuclara sahip olabilir Bu bozukluk LDL icin fonksiyonel reseptorlerin yoklugu ile karakterizedir LDL reseptorlerinin olusumundaki eksiklikler endositik yolu bozan LDL nin karacigere ve diger hucrelere girisini engelleyen hatali reseptorlere neden olabilir Bu kan plazmasinda LDL birikmesine neden olur bu da arterleri daraltan ve kalp krizi riskini artiran aterosklerotik plaklara neden olur Bu genetik hastalik kendini yaralama zihinsel yetersizlik ve gut ile karakterizedir Purin nukleotid olusumu icin gerekli bir enzim olan yoklugundan kaynaklanir Enzim eksikligi gerekli nukleotidlerin seviyesini azaltir ve biyosentez ara urunlerinin birikmesine neden olur bu da yukarida belirtilen olagandisi davranislarla sonuclanir SCID SCID T hucrelerinin kaybi ile karakterizedir Bu bagisiklik sistemi bilesenlerinin eksikligi etkilenen bireyler immunolojik hafiza gelistiremediginden bulasici ajanlara duyarliligi arttirir Bu immunolojik bozukluk dATP birikmesine neden olan aktivitesindeki bir eksiklikten kaynaklanir Bu dATP molekulleri daha sonra DNA sentezini onleyen ribonukleotid reduktazi inhibe eder Huntington hastaligi Bu norolojik hastalik DNA sentezi sirasinda meydana gelen hatalardan kaynaklanir Bu hatalar veya mutasyonlar gendeki genisleyen CAG trinukleotit tekrarlari tarafindan kodlanan tekrarlayan glutamin tortulari iceren bir mutant Huntingtin proteininin ekspresyonuna yol acar Huntington hastaligi noronal kayip ve gliozis ile karakterizedir Hastaligin belirtileri hareket bozuklugu bilissel gerileme ve davranis bozuklugudur Ayrica bakinizLipit Hucre zari Nukleotit DNA DNA replikasyonu Prostaglandin Porfirin B12 vitamini EnzimKaynakca a b c d Alberts Bruce 2008 Molecular Biology of the Cell 5 bas New York Garland Science ISBN 978 0815341055 Zumdahl Steven S Zumdahl Susan A 2008 Chemistry 8 bas CA Cengage Learning ISBN 978 0547125329 Voet Donald Voet Judith G Pratt Charlotte W 2013 Fundamentals of Biochemistry Life at the Molecular Level 4th Hoboken NJ Wiley ISBN 978 0470547847 a b c d e Lodish Harvey 2007 Molecular cell biology 6 New York W H Freeman ISBN 978 0716743668 a b c d e Cox David L Nelson Michael M 2008 Lehninger principles of biochemistry 5 bas New York W H Freeman ISBN 9780716771081 Hanin Israel 2013 Phospholipids Biochemical Pharmaceutical and Analytical Considerations Springer ISBN 978 1475713664 a b c d Vance Dennis E Vance Jean E 2008 Biochemistry of lipids lipoproteins and membranes 5 bas Amsterdam Elsevier ISBN 978 0444532190 Katsaras J 2001 Lipid bilayers structure and interactions with 6 tables Berlinisbn 978 3540675556 Springer a b c d e Stryer Jeremy M Berg John L Tymoczko Lubert 2007 Biochemistry 6 bas New York Freeman ISBN 978 0716787242 Gault CR LM Obeid YA Hannun 2010 An Overview of sphingolipid metabolism from synthesis to breakdown Advances in Experimental Medicine and Biology 688 s 1 23 doi 10 1007 978 1 4419 6741 1 1 ISBN 978 1 4419 6740 4 PMC 3069696 2 PMID 20919643 a b Siegel George J 1999 Basic neurochemistry molecular cellular and medical aspects 6 bas Philadelphia Pa u a Lippincott Williams amp Wilkins ISBN 978 0397518203 a b c Harris J Robin 2010 Cholesterol binding and cholesterol transport proteins structure and function in health and disease Dordrecht Springer ISBN 978 9048186211 a b c d e f Watson James D 2007 Molecular biology of the gene 6 bas San Francisco Calif Benjamin Cummings ISBN 978 0805395921 Kappock TJ Ealick SE Stubbe J Ekim 2000 Modular evolution of the purine biosynthetic pathway Current Opinion in Chemical Biology 4 5 s 567 572 doi 10 1016 s1367 5931 00 00133 2 PMID 11006546 Sampei G Baba S Kanagawa M Yanai H Ishii T Kawai H Fukai Y Ebihara A Nakagawa N Kawai G Ekim 2010 Crystal structures of glycinamide ribonucleotide synthetase PurD from thermophilic eubacteria Journal of Biochemistry 148 4 s 429 438 doi 10 1093 jb mvq088 PMID 20716513 erisim tarihi kullanmak icin url gerekiyor yardim Hoskins AA Anand R Ealick SE Stubbe J 17 Agustos 2004 The formylglycinamide ribonucleotide amidotransferase complex from Bacillus subtilis metabolite mediated complex formation Biochemistry 32 43 bas s 10314 27 doi 10 1021 bi049127h PMID 15301530 erisim tarihi kullanmak icin url gerekiyor yardim Mueller EJ Meyer E Rudolph J Davisson VJ Stubbe J 1 Mart 1994 N5 carboxyaminoimidazole ribonucleotide evidence for a new intermediate and two new enzymatic activities in the de novo purine biosynthetic pathway of Escherichia coli Biochemistry 8 33 bas s 2269 78 doi 10 1021 bi00174a038 PMID 8117684 erisim tarihi kullanmak icin url gerekiyor yardim Firestine SM Poon SW Mueller EJ Stubbe J Davisson VJ 4 Ekim 1994 Reactions catalyzed by 5 aminoimidazole ribonucleotide carboxylases from Escherichia coli and Gallus gallus a case for divergent catalytic mechanisms Biochemistry 39 33 bas ss 11927 34 doi 10 1021 bi00205a031 PMID 7918411 erisim tarihi kullanmak icin url gerekiyor yardim a b Srere PA 1987 Complexes of sequential metabolic enzymes Annual Review of Biochemistry 1 56 bas s 89 124 doi 10 1146 annurev bi 56 070187 000513 PMID 2441660 Broach edited by Jeffrey N Strathern Elizabeth W Jones James R 1981 The Molecular biology of the yeast Saccharomyces Cold Spring Harbor N Y Cold Spring Harbor Laboratory ISBN 978 0879691394 KB1 bakim Fazladan yazi yazar listesi link a b O Donovan GA Neuhard J Eylul 1970 Pyrimidine metabolism in microorganisms Bacteriological Reviews 3 34 bas s 278 343 doi 10 1128 MMBR 34 3 278 343 1970 PMC 378357 2 PMID 4919542 a b c d Geer Gerald Karp responsible for the revision of chapter 15 Peter van der 2004 Cell and molecular biology concepts and experiments 4 Wiley International bas New York J Wiley amp Sons ISBN 978 0471656654 a b Griffiths Anthony J F 1999 Modern genetic analysis 2 bas New York Freeman ISBN 978 0716731184 a b Wu G Mayis 2009 Amino acids metabolism functions and nutrition Amino Acids 1 s 1 17 doi 10 1007 s00726 009 0269 0 PMID 19301095 erisim tarihi kullanmak icin url gerekiyor yardim Mousdale D M Coggins J R 1991 Amino Acid Synthesis Target Sites for Herbicide Action s 29 56 doi 10 1007 978 1 4899 2433 9 2 ISBN 978 1 4899 2435 3 Miflin B J Lea P J 1977 Amino Acid Metabolism Annual Review of Plant Physiology 28 s 299 329 doi 10 1146 annurev pp 28 060177 001503 a b c Umbarger HE 1978 Amino acid biosynthesis and its regulation Annual Review of Biochemistry 47 1 s 532 606 doi 10 1146 annurev bi 47 070178 002533 PMID 354503 Perez Arellano I Carmona Alvarez F Martinez AI Rodriguez Diaz J Cervera J Mart 2010 Pyrroline 5 carboxylate synthase and proline biosynthesis from osmotolerance to rare metabolic disease Protein Science 19 3 s 372 82 doi 10 1002 pro 340 PMC 2866264 2 PMID 20091669 erisim tarihi kullanmak icin url gerekiyor yardim Xu Y Labedan B Glansdorff N Mart 2007 Surprising arginine biosynthesis a reappraisal of the enzymology and evolution of the pathway in microorganisms Microbiology and Molecular Biology Reviews 71 1 s 36 47 doi 10 1128 MMBR 00032 06 PMC 1847373 2 PMID 17347518 MetaCyc L lysine biosynthesis I 4 Mayis 2022 tarihinde kaynagindan Erisim tarihi 15 Nisan 2022 PETERKOFSKY B GILVARG C Mayis 1961 N Succinyl L diaminopimelic glutamic transaminase The Journal of Biological Chemistry 236 5 s 1432 8 doi 10 1016 S0021 9258 18 64192 4 PMID 13734750 KINDLER SH GILVARG C Aralik 1960 N Succinyl L 2 6 diaminopimelic acid deacylase The Journal of Biological Chemistry Cilt 235 s 3532 3535 doi 10 1016 S0021 9258 18 64502 8 PMID 13756049 Born TL Blanchard JS Ekim 1999 Structure function studies on enzymes in the diaminopimelate pathway of bacterial cell wall biosynthesis Current Opinion in Chemical Biology 3 5 s 607 613 doi 10 1016 s1367 5931 99 00016 2 PMID 10508663 Escherichia coli K 12 substr MG1655 serine biosynthesis SRI International 15 Agustos 2016 tarihinde kaynagindan Erisim tarihi 12 Aralik 2013 Bell JK Grant GA Banaszak LJ 30 Mart 2004 Multiconformational states in phosphoglycerate dehydrogenase Biochemistry 43 12 s 3450 8 doi 10 1021 bi035462e PMID 15035616 erisim tarihi kullanmak icin url gerekiyor yardim Dubnovitsky AP Kapetaniou EG Papageorgiou AC Ocak 2005 Enzyme adaptation to alkaline pH atomic resolution 1 08 A structure of phosphoserine aminotransferase from Bacillus alcalophilus Protein Science 14 1 s 97 110 doi 10 1110 ps 041029805 PMC 2253317 2 PMID 15608117 Wang W Kim R Jancarik J Yokota H Kim SH 10 Ocak 2001 Crystal structure of phosphoserine phosphatase from Methanococcus jannaschii a hyperthermophile at 1 8 A resolution Structure 9 1 s 65 71 doi 10 1016 s0969 2126 00 00558 x PMID 11342136 Monschau N Stahmann KP Sahm H McNeil JB Bognar AL 1 Mayis 1997 Identification of Saccharomyces cerevisiae GLY1 as a threonine aldolase a key enzyme in glycine biosynthesis FEMS Microbiology Letters 150 1 s 55 60 doi 10 1111 j 1574 6968 1997 tb10349 x PMID 9163906 Pye VE Tingey AP Robson RL Moody PC 24 Eylul 2004 The structure and mechanism of serine acetyltransferase from Escherichia coli The Journal of Biological Chemistry 279 39 ss 40729 36 doi 10 1074 jbc M403751200 PMID 15231846 Huang B Vetting MW Roderick SL Mayis 2005 The active site of O acetylserine sulfhydrylase is the anchor point for bienzyme complex formation with serine acetyltransferase Journal of Bacteriology 187 9 s 3201 5 doi 10 1128 JB 187 9 3201 3205 2005 PMC 1082839 2 PMID 15838047 McPhalen CA Vincent MG Picot D Lesk AM Chothia C 5 Eylul 1992 Domain closure in mitochondrial aspartate aminotransferase Journal of Molecular Biology 227 1 s 197 213 doi 10 1016 0022 2836 92 90691 C PMID 1522585 Larsen TM Boehlein SK Schuster SM Richards NG Thoden JB Holden HM Rayment I 7 Aralik 1999 Three dimensional structure of Escherichia coli asparagine synthetase B a short journey from substrate to product Biochemistry 38 49 ss 16146 57 CiteSeerX 10 1 1 453 5998 2 doi 10 1021 bi9915768 PMID 10587437 Velasco AM Leguina JI Lazcano A October 2002 Molecular evolution of the lysine biosynthetic pathways Journal of Molecular Evolution 55 4 ss 445 59 Bibcode 2002JMolE 55 445V doi 10 1007 s00239 002 2340 2 PMID 12355264 Kotaka M Ren J Lockyer M Hawkins AR Stammers DK 20 Ekim 2006 Structures of R and T state Escherichia coli aspartokinase III Mechanisms of the allosteric transition and inhibition by lysine The Journal of Biological Chemistry 281 42 ss 31544 52 doi 10 1074 jbc M605886200 PMID 16905770 Hadfield A Kryger G Ouyang J Petsko GA Ringe D Viola R 18 Haziran 1999 Structure of aspartate beta semialdehyde dehydrogenase from Escherichia coli a key enzyme in the aspartate family of amino acid biosynthesis Journal of Molecular Biology 289 4 ss 991 1002 doi 10 1006 jmbi 1999 2828 PMID 10369777 Mirwaldt C Korndorfer I Huber R 10 Subat 1995 The crystal structure of dihydrodipicolinate synthase from Escherichia coli at 2 5 A resolution Journal of Molecular Biology 246 1 ss 227 39 doi 10 1006 jmbi 1994 0078 PMID 7853400 Cirilli M Zheng R Scapin G Blanchard JS 16 Eylul 2003 The three dimensional structures of the Mycobacterium tuberculosis dihydrodipicolinate reductase NADH 2 6 PDC and NADPH 2 6 PDC complexes Structural and mutagenic analysis of relaxed nucleotide specificity Biochemistry 42 36 ss 10644 50 doi 10 1021 bi030044v PMID 12962488 Beaman TW Binder DA Blanchard JS Roderick SL 21 Ocak 1997 Three dimensional structure of tetrahydrodipicolinate N succinyltransferase Biochemistry 36 3 ss 489 94 doi 10 1021 bi962522q PMID 9012664 Weyand S Kefala G Weiss MS 30 Mart 2007 The three dimensional structure of N succinyldiaminopimelate aminotransferase from Mycobacterium tuberculosis Journal of Molecular Biology 367 3 ss 825 38 doi 10 1016 j jmb 2007 01 023 PMID 17292400 Nocek BP Gillner DM Fan Y Holz RC Joachimiak A 2 Nisan 2010 Structural basis for catalysis by the mono and dimetalated forms of the dapE encoded N succinyl L L diaminopimelic acid desuccinylase Journal of Molecular Biology 397 3 ss 617 26 doi 10 1016 j jmb 2010 01 062 PMC 2885003 2 PMID 20138056 Pillai B Cherney M Diaper CM Sutherland A Blanchard JS Vederas JC James MN 23 Kasim 2007 Dynamics of catalysis revealed from the crystal structures of mutants of diaminopimelate epimerase Biochemical and Biophysical Research Communications 363 3 ss 547 53 doi 10 1016 j bbrc 2007 09 012 PMID 17889830 Gokulan K Rupp B Pavelka MS Jr Jacobs WR Jr Sacchettini JC 16 Mayis 2003 Crystal structure of Mycobacterium tuberculosis diaminopimelate decarboxylase an essential enzyme in bacterial lysine biosynthesis The Journal of Biological Chemistry 278 20 ss 18588 96 doi 10 1074 jbc M301549200 PMID 12637582 a b c Weaver Robert F 2005 Molecular biology 3 bas Boston McGraw Hill Higher Education ISBN 978 0 07 284611 9 a b c d e Cooper Geoffrey M 2000 The Cell A Molecular Approach 2 bas Washington DC ASM Press ISBN 978 0878931064 Jackson R J Subat 2010 The mechanism of eukaryotic translation initiation and principles of its regulation Molecular Cell Biology 10 2 ss 113 127 doi 10 1038 nrm2838 PMC 4461372 2 PMID 20094052 Green Rachel Harry F Noller 1997 Ribosomes and Translation Annu Rev Biochem Cilt 66 ss 679 716 doi 10 1146 annurev biochem 66 1 679 PMID 9242921 a b c d Weissbach Herbert Pestka Sidney 1977 Molecular Mechanisms of Protein Biosynthesis New York Academic Press ISBN 978 0127442501 Frank J Haixiao Gao Eylul 2007 The process of mRNA tRNA translocation PNAS 104 50 ss 19671 19678 doi 10 1073 pnas 0708517104 PMC 2148355 2 PMID 18003906 a b c Bandeali Salman J Daye Jad Virani Salim S 30 Kasim 2013 Novel Therapies for Treating Familial Hypercholesterolemia Current Atherosclerosis Reports 16 1 s 382 doi 10 1007 s11883 013 0382 0 PMID 24293346 a b c Kang Tae Hyuk Park Yongjin Bader Joel S Friedmann Theodore Cooney Austin John 9 Ekim 2013 The Housekeeping Gene Hypoxanthine Guanine Phosphoribosyltransferase HPRT Regulates Multiple Developmental and Metabolic Pathways of Murine Embryonic Stem Cell Neuronal Differentiation PLOS ONE 8 10 ss e74967 Bibcode 2013PLoSO 874967K doi 10 1371 journal pone 0074967 PMC 3794013 2 PMID 24130677 a b c Walport Ken Murphy Paul Travers Mark 2011 Janeway s Immunobiology 8 Oxford Taylor amp Francis ISBN 978 0815342434 a b Hughes edited by Donald C Lo Robert E 2010 Neurobiology of Huntington s disease applications to drug discovery 2nd Boca Raton CRC Press Taylor amp Francis Group ISBN 978 0849390005 KB1 bakim Fazladan yazi yazar listesi link Biglan Kevin M Ross Christopher A Langbehn Douglas R Aylward Elizabeth H Stout Julie C Queller Sarah Carlozzi Noelle E Duff Kevin Beglinger Leigh J Paulsen Jane S 26 Haziran 2009 Motor abnormalities in premanifest persons with Huntington s disease The PREDICT HD study Movement Disorders 24 12 ss 1763 1772 doi 10 1002 mds 22601 PMC 3048804 2 PMID 19562761